Раздел 6. Надорганизменные системы. Эволюция органического мира
 Скачать 10.97 Mb. Скачать 10.97 Mb.
|

|
Усложнение живых организмов в процессе эволюции Историю органического мира на Земле изучают по сохранившимся остаткам, отпечаткам и другим следам жизнедеятельности живых организмов. Она является предметом науки палеон тологии. Исходя из того, что остатки разных организмов расположены в различных пластах гор ных пород, была создана геохронологическая шкала, согласно которой историю Земли разделили на определенные промежутки времени: зоны, эры, периоды и века (табл. 6.1). Эоном называют большой промежуток времени в геологической истории, объединяющий не сколько эр. В настоящее время выделяют только два зона: криптозой (скрытая жизнь) и фанеро- зой (явная жизнь). Эра — это промежуток времени в геологической истории, являющийся подраз делением эона, объединяющий, в свою очередь, периоды. В криптозое выделяют две эры (архей и протерозой), тогда как в фанерозое — три (палеозой, мезозой и кайнозой). Важную роль в создании геохронологической шкалы сыграли руководящие ископаемые — остатки организмов, которые были многочисленны в определенные промежутки времени и хоро шо сохранились. Развитие жизни в криптозое. Архей и протерозой составляют большую часть истории жизни (период 4,6 млрд лет — 0,6 млрд лет назад), однако сведений о жизни в тот период недоста точно. Первые остатки органических веществ биогенного происхождения имеют возраст около 3,8 млрд лет, а прокариотические организмы существовали уже 3,5 млрд лет назад. Первые прокариоты входили в состав специфических экосистем — цианобактериальных матов, благо даря деятельности которых образовались специфические осадочные породы строматолиты («ка менные ковры»). Понять жизнь давних прокариотических экосистем помогло открытие их современных анало гов — строматолитов в заливе Шарк-Бей в Австралии и специфических пленок на поверхности почвы в заливе Сиваш в Украине. На поверхности цианобактериальных матов расположены фото- синтезирующие цианобактерии, а под их слоем — чрезвычайно разнообразные бактерии других групп и архей. Минеральные вещества, которые оседают на поверхность мата и образовываются за счет его жизнедеятельности, откладываются пластами (приблизительно 0,3 мм в год). Такие примитивные экосистемы могут существовать лишь в непригодных для жизни других организмов местах, и действительно, оба вышеупомянутые местообитания характеризуются чрезвычайно вы сокой соленостью. Многочисленные данные свидетельствуют о том, что поначалу Земля имела атмосферу воз обновляемого характера, в состав которой входили: углекислый газ, водяной пар, оксид серы, а также угарный газ, водород, сероводород, аммиак, метан и т. п. Первые организмы Земли были анаэробами, однако благодаря фотосинтезу цианобактерий в среду выделялся свободный кисло род, который сначала быстро связывался с восстановителями, находящимися в среде, и лишь по сле связывания всех восстановителей среда начала приобретать окислительные свойства. О таком переходе свидетельствуют отложение окисленных форм железа — гематита и магнетита. Около 2 млрд лет назад в результате геофизических процессов практически все несвязанное в осадочных породах железо переместилось к ядру планеты, а кислород начал накапливаться в ат мосфере из-за отсутствия этого элемента — произошла «кислородная революция». Она явилась переломным этапом в истории Земли, который повлек за собой не только смену состава атмосфе ры и образование озонового экрана в атмосфере — главной предпосылки для заселения суши, но и состава пород, формирующихся на поверхности Земли. В протерозое произошло и другое важное событие — возникновение эукариот. В последние годы удалось собрать убедительные доказательства теории эндосимбиогенетического происхожде ния эукариотической клетки — путем симбиоза нескольких прокариотических клеток. Вероятно, «главным» предком эукариот стали архей, которые перешли к поглощению пищевых частиц пу тем фагоцитоза. Наследственный аппарат переместился вглубь клетки, сохранив, тем не менее, связь с мембраной благодаря переходу внешней мембраны возникшей ядерной оболочки в мембра ны эндоплазматической сети. 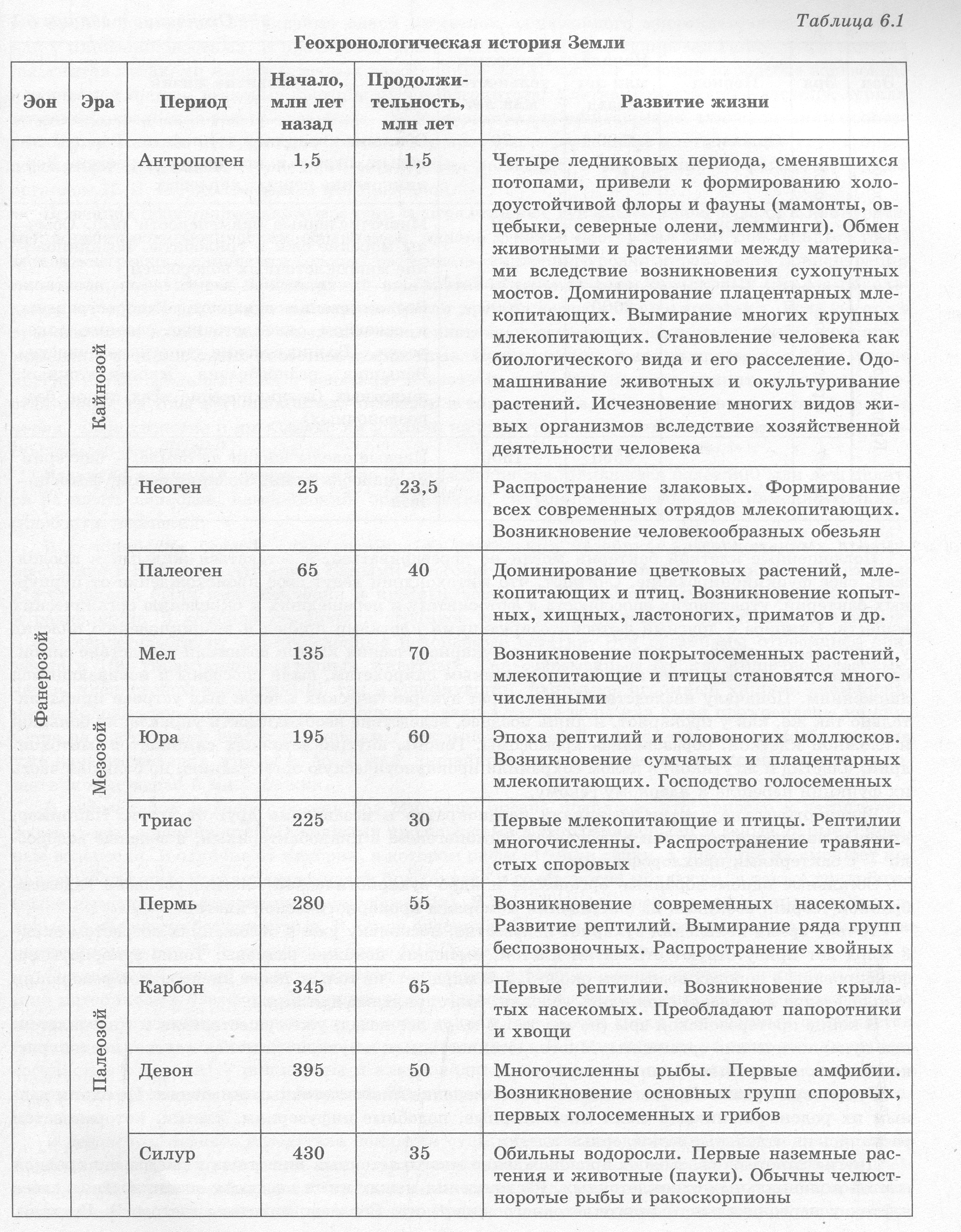 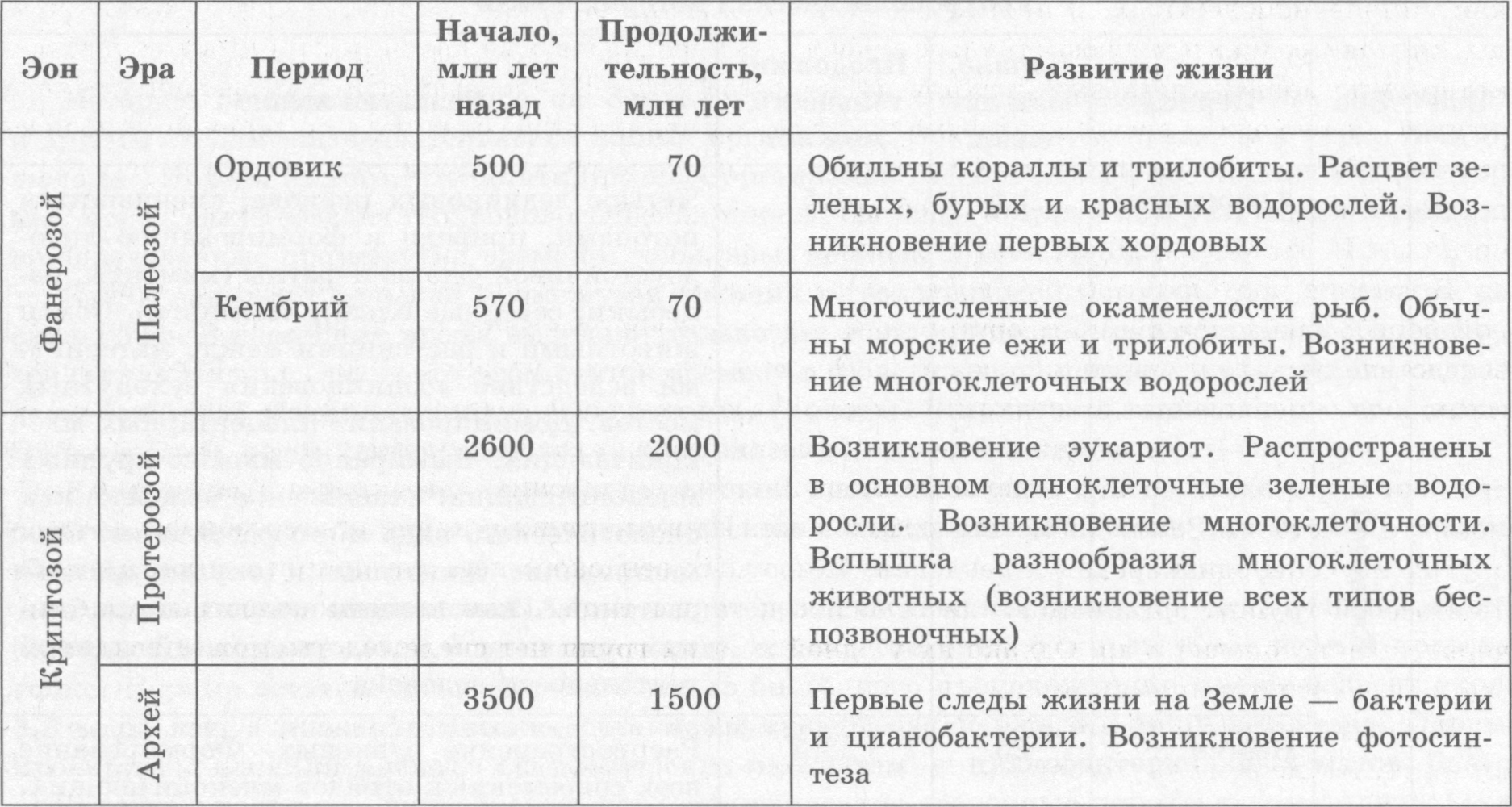 Поглощенные клеткой бактерии могли не перевариваться, а оставаться живыми и продол жать свое функционирование. Считают, что митохондрии ведут свое происхождение от пурпур ных бактерий, утративших способность к фотосинтезу и перешедших к окислению органических веществ. Симбиоз с другими фотосинтезирующими клетками привел к возникновению пластид у растительных клеток. Вероятно, жгутики эукариотических клеток возникли вследствие симби оза с бактериями, которые, подобно современным спирохетам, были способны к извивающимся движениям. Поначалу наследственный аппарат эукариотических клеток был устроен приблизи тельно так же, как у прокариот, и лишь позднее, вследствие необходимости управления большой и сложной клеткой, образовались хромосомы. Геномы внутриклеточных симбионтов (митохон дрий, пластид и жгутиков) в целом сохранили прокариотическую организацию, но большая часть их функций перешла к ядерному геному. Эукариотические клетки возникали неоднократно и независимо друг от друга. Например, красные водоросли возникли в результате симбиогенеза с цианобактериями, а зеленые водорос ли — с бактериями-прохлорофитами. Остальные одномембранные органеллы и ядро эукариотической клетки, согласно эндомем- бранной теории, возникли из впячиваний мембраны прокариотической клетки. Точное время появления эукариот неизвестно, поскольку уже в отложениях возрастом около 3 млрд лет присутствуют отпечатки клеток, имеющих похожие размеры. Точно эукариоты за фиксированы в породах возрастом около 1,5-2 млрд лет, но только после кислородной революции (около 1 млрд лет назад) сложились условия, благоприятные для них. В конце протерозойской эры (не менее 1,5 млрд лет назад) уже существовали и многоклеточ ные эукариотические организмы. Многоклеточность, как и эукариотическая клетка, неоднократ но возникала у разных групп организмов. Существуют различные взгляды на происхождение многоклеточных животных. По одним дан ным их родоначальниками были многоядерные, подобные инфузориям, клетки, которые затем распались на отдельные одноядерные клетки. Другие гипотезы связывают происхождение многоклеточных животных с дифференцировкой клеток колониальных одноклеточных. Расхождения между ними касаются возникновения слоев клеток у первоначального многоклеточного животного. Согласно гипотезы гастреи Э. Геккеля, то происходит путем впячивания одной из стенок однослойного многоклеточного организма, как у кишечнополостных. В противовес ей И. И. Мечников сформулировал гипотезу фагоцител- лы, считая предками многоклеточных однослойные шарообразные колонии наподобие вольвокса, которые поглощали пищевые частицы путем фагоцитоза. Клетка, захватившая частицу, теряла жгутик и переходила вглубь организма, где и осуществляла пищеварение, а по окончании процес са возвращалась на поверхность. Со временем произошло разделение клеток на два слоя с опре деленными функциями — внешний обеспечивал движение, а внутренний — фагоцитоз. Такой организм И. И. Мечников назвал фагоцителлой. В течение продолжительного времени многоклеточные эукариоты проигрывали в конкурент ной борьбе прокариотическим организмам, однако в конце протерозоя (800-600 млн лет тому) вследствие резкого изменения условий на Земле — снижения уровня морей, роста концентрации кислорода, уменьшения концентрации карбонатов в морской воде, регулярных циклов похоло дания — многоклеточные эукариоты получили преимущества над прокариотами. Если до этого времени встречались только отдельные многоклеточные растения и, возможно, грибы, то с этого момента в истории Земли известны и животные. Из возникших в конце протерозоя фаун лучше других изучены эдиакарская и вендская. Животных вендского периода принято включать в со став особой группы организмов или относить к таким типам, как кишечнополостные, плоские черви, членистоногие и др. Однако ни у одной из этих групп нет скелетов, что может свидетель ствовать об отсутствии хищников. Развитие жизни в палеозойской эре. Палеозойская эра, длившаяся более 300 млн лет, делит ся на шесть периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный (карбон) и пермский. В кембрийском периоде суша состояла из нескольких материков, расположенных преиму щественно в Южном полушарии. Самыми многочисленными фотосинтезирующими организмами в этот период были цианобактерии и красные водоросли. В толще воды жили фораминиферы и радиолярии. В кембрии появляется огромное количество скелетных животных организмов, о чем свидетельствуют многочисленные ископаемые остатки. Эти организмы относились при мерно к 100 типам многоклеточных животных, как современным (губки, кишечнополостные, черви, членистоногие, моллюски), так и исчезнувшим, например: огромный хищник аномалока- рис и колониальные граптолиты, которые плавали в толще воды или были прикреплены ко дну. Суша на протяжении кембрия оставалась почти незаселенной, однако процесс почвообразования уже начали бактерии, грибы и, возможно, лишайники, а в конце периода на сушу вышли мало- щетинковые черви и многоножки. В ордовикском периоде уровень вод Мирового океана поднялся, что привело к затоплению материковых низменностей. Основными продуцентами в этот период были зеленые, бурые и крас ные водоросли. В отличие от кембрия, в котором рифы строили губки, в ордовике их сменяют ко ралловые полипы. Расцвет переживали брюхоногие и головоногие моллюски, а также трилобиты (ныне вымершие родственники паукообразных). В этом периоде впервые зафиксированы и хор довые, в частности бесчелюстные. В конце ордовика произошло грандиозное вымирание, которое уничтожило около 35 % семейств и более 50 % родов морских животных. Силурийский период характеризуется усилением горообразования, которое привело к осуше нию материковых платформ. Ведущую роль в фауне беспозвоночных силура играли головоногие моллюски, иглокожие и гигантские ракоскорпионы, тогда как среди позвоночных сохраняется большое разнообразие бесчелюстных и появляются рыбы. В конце периода на сушу вышли первые сосудистые растения — риниофиты и плауновидные, которые начали колонизацию мелководья и приливно-отливной зоны побережий. На сушу вышли и первые представители класса пауко образных. В девонском периоде вследствие поднятия суши образовались большие мелководья, которые пересыхали и даже промерзали, поскольку климат становился еще более континентальным, чем в силуре. В морях преобладают кораллы и иглокожие, тогда как головоногие моллюски представ лены спирально закрученными аммонитами. Среди позвоночных девона расцвета достигли рыбы, причем на смену панцирным пришли и хрящевые, и костные, а также двоякодышащие и кисте- перые. В конце периода появляются первые амфибии, которые сначала жили в воде. В среднем девоне на суше появились первые леса из папоротников, плаунов и хвощей, которые были заселены червями и многочисленными членистоногими (многоножками, пауками, скорпио нами, бескрылыми насекомыми). В конце девона появились первые голосеменные. Освоение суши растениями привело к уменьшению выветривания и усилению почвообразования. Закрепление почв привело к возникновению русел рек. В каменноугольном периоде суша была представлена двумя материками, разделенными океа ном, а климат стал заметно более теплым и влажным. К концу периода произошло небольшое под нятие суши, а климат сменился более континентальным. В морях господствовали фораминиферы, кораллы, иглокожие, хрящевые и костные рыбы, а пресные водоемы населяли двухстворчатые моллюски, ракообразные и разнообразные земноводные. В середине карбона возникли мелкие на секомоядные рептилии, а среди насекомых появились крылатые (тараканы, стрекозы). Для тропиков были характерны заболоченные леса, в которых доминировали гигантские хво щи, плауны и папоротники, отмершие остатки которых образовали впоследствии залежи камен ного угля. В середине периода в умеренной зоне, благодаря их независимости от воды в процессе оплодотворения и наличию семени, началось распространение голосеменных. Пермский период отличался слиянием всех материков в единый суперконтинент Пангею, от ступлением морей и усилением континентальности климата до такой степени, что во внутренних районах Пангеи образовались пустыни. К концу периода на суше почти исчезли древовидные папоротники, хвощи и плауны, а господствующее положение заняли засухоустойчивые голосе менные. Несмотря на то, что крупные амфибии еще продолжали существовать, возникли разные груп пы рептилий, в том числе крупных растительноядных и хищных. В конце перми произошло са мое большое вымирание в истории жизни, так как исчезли многие группы кораллов, трилобиты, большинство головоногих, рыб (в первую очередь хрящевых и кистеперых), а также амфибий. Морская фауна потеряла при этом 40-50% семейств и около 70% родов. Развитие жизни в мезозое. Мезозойская эра продолжалась около 165 млн лет и характеризо валась поднятием суши, интенсивным горообразованием и снижением влажности климата. Она делится на три периода: триасовый, юрский и меловой. В начале триасового периода климат был засушливым, однако позднее вследствие поднятия уровня морей он стал более влажным. Среди растений преобладали голосеменные, папоротники и хвощи, однако древесные формы споровых практически полностью вымерли. Высокого разви тия достигли некоторые кораллы, аммониты, новые группы фораминифер, двухстворчатых мол люсков и иглокожих, тогда как разнообразие хрящевых рыб уменьшилось, изменились и группы костных рыб. Господствовавшие на суше рептилии начали осваивать и водную среду, как ихтио завры и плезиозавры. Из пресмыкающихся триаса до нашего времени дожили крокодилы, гатте- рии и черепахи. В конце триаса появились динозавры, млекопитающие и птицы. В юрском периоде суперконтинент Пангея раскололся на несколько меньших. Большая часть юры была очень влажной, а к его концу климат стал более засушливым. Доминирующей группой растений были голосеменные, из которых от того времени сохранились секвойи. В морях про цветали моллюски (аммониты и белемниты, двухстворчатые и брюхоногие), губки, морские ежи, хрящевые и костные рыбы. Крупные амфибии практически полностью вымерли в юрском перио де, однако появились современные группы земноводных (хвостатые и бесхвостые) и чешуйчатых (ящериц и змей), возросло разнообразие млекопитающих. К концу периода возникли и возмож ные предки первых птиц — археоптериксы. Однако во всех экосистемах доминировали пресмыка ющиеся — ихтиозавры и плезиозавры, динозавры и летающие ящеры — птерозавры. Меловой период получил название в связи с образованием мела в осадочных породах того времени. На всей Земле, кроме приполярных областей, был стойкий теплый и влажный климат. В этом периоде возникли и приобрели широкое распространение покрытосеменные, вытеснявшие голосеменных, что повлекло за собой резкое увеличение разнообразия насекомых. В морях, по мимо моллюсков, костистых рыб, плезиозавров, вновь появилось огромное количество форами-нифер, раковинки которых и образовали залежи мела, а на суше преобладали динозавры. Лучше приспособленные к воздушной среде птицы начали постепенно вытеснять летающих ящеров. В конце периода произошло глобальное вымирание, в результате которого исчезли аммониты, белемниты, динозавры, птерозавры и морские ящеры, древние группы птиц, а также некоторые голосеменные. С лица Земли в целом исчезло около 16% семейств и 50% родов животных. Кри зис в конце мела связывают с падением большого метеорита в Мексиканский залив, однако он, скорее всего, не был единственной причиной глобальных изменений. В ходе последующего похо лодания выжили только небольшие рептилии и теплокровные млекопитающие. Развитие жизни в кайнозое. Кайнозойская эра началась около 66 млн лет назад и продолжа ется до настоящего времени. Она характеризуется господством насекомых, птиц, млекопитающих и покрытосеменных растений. Кайнозой делят на три периода — палеоген, неоген и антропо- ген — последний из которых является самым коротким в истории Земли. В раннем и среднем палеогене климат оставался теплым и влажным, к концу периода стало прохладнее и суше. Доминирующей группой растений стали покрытосеменные, однако, если в на чале периода преобладали вечнозеленые леса, то в конце появилось много листопадных, а в за сушливых зонах образовались степи. Среди рыб господствующее положение заняли костистые рыбы, а количество видов хрящевых, несмотря на их заметную роль в соленых водоемах, незначительно. На суше из рептилий сохра нились только чешуйчатые, крокодилы и черепахи, тогда как млекопитающие заняли большую часть их экологических ниш. В середине периода появились основные отряды млекопитающих, в том числе насекомоядные, хищные, ластоногие, китообразные, копытные и приматы. Изоляция материков сделала фауну и флору географически более разнообразными: Южная Америка и Ав стралия стали центрами развития сумчатых, а другие материки — плацентарных млекопитающих. Неогеновый период. Земная поверхность в неогене приобрела современный вид. Климат стал более прохладным и сухим. В неогене уже сформировались все отряды современных млекопита ющих, а в африканских саванах возникло семейство Гоминид и род Человек. К концу периода в приполярных областях континентов распространились хвойные леса, появились тундры, а сте пи умеренного пояса заняли злаки. Четвертичный период (антропоген) характеризуется периодическими сменами оледенений и потеплений. Во время оледенений высокие широты покрывались ледниками, резко снижался уровень океана, суживались тропический и субтропический пояса. На близлежащих к ледникам территориях устанавливался холодный и сухой климат, который способствовал формированию холодоустойчивых групп животных — мамонтов, гигантских оленей, пещерных львов и др. Со путствовавшее процессу оледенения снижение уровня Мирового океана привело к образованию сухопутных мостов между Азией и Северной Америкой, Европой и Британскими островами и т. д. Миграции животных, с одной стороны, привели к взаимообогащению флор и фаун, а с другой, к вытеснению реликтов пришельцами, например, сумчатых и копытных в Южной Америке. Эти процессы, однако, не затронули Австралию, оставшуюся изолированной. В целом, периодические изменения климата привели к формированию чрезвычайно обильного видового разнообразия, характерного для нынешнего этапа эволюции биосферы, а также оказали влияние на эволюцию человека. На протяжении антропогена несколько видов рода Человек рас селились из Африки в Евразию. Около 200 тысяч лет назад в Африке возник вид Человек разум ный, который после продолжительного периода существования в Африке около 70 тысяч лет на зад вышел в Евразию и около 35-40 тыс. лет назад — в Америку. После периода сосуществования с близкородственными видами он вытеснил их и расселился по всей территории земного шара. Около 10 тыс. лет назад хозяйственная деятельность человека в умеренно теплых областях земного шара начала оказывать влияние как на облик планеты (распашка земель, выжигание лесов, перевыпас пастбищ, опустынивание и т. д.), так и на животный и растительный мир вследствие сокращения ареалов их обитания и истребления, и вступил в действие антропогенный фактор. |