термодинамика реферат. Основные понятия и исходные положения термодинамики
 Скачать 141.71 Kb. Скачать 141.71 Kb.
|

|
ГЛАВА 3 АНАЛИТИЧЕСКИЕ И ЧИСЛЕННЫЕ ИССЛЕДОВАНИЯ САМООРГАНИЗАЦИИ РАЗЛИЧНЫХ СИСТЕМ. 3.1. ЯЧЕЙКИ БЕНАРА . Для того , чтобы экспериментально изучить структуры , достаточно иметь сковороду , немного масла и какой ни будь мелкий порошок , чтобы было заметно движение жидкости . Нальем в сковороду масло с размешанным в нем порошком и будем подогревать ее снизу (рис. 3.1) 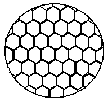 Рис. 3.1. Конвективные ячейки Бенара. Если дно сковороды плоское и нагреваем мы ее равномерно , то можно считать , что у дна и на поверхности поддерживаются постоянные температуры , снизу - Т1 , сверху - Т2 . Пока разность температуры Т = Т1 - Т2 невелика , частички порошка неподвижны , а следовательно , неподвижна и жидкость . Будем плавно увеличивать температуру Т1 . С ростом разности температур до значения Тc наблюдается все та же картина , но когда Т Тc , вся среда разбивается на правильные шестигранные ячейки (см. Рис. 3.1) в центре каждой из которых жидкость движется вверх , по кроям вниз . Если взять другую сковороду , то можно убедиться , что величина возникающих ячеек практически не зависит от ее формы и размеров . Этот замечательный опыт впервые был проделан Бенаром в начале нашего века , а сами ячейки получили название ячеек Бенара . Элементарное качественное объяснения причины движения жидкости заключается в следующем . Из-за теплового расширения жидкость расслаивается , и в более нижнем слое плотность жидкости 1 меньше , чем в верхнем 2 . Возникает инверсный градиент плотности , направленный противоположно силе тяжести . Если выделить элементарный объем V , который немного смещается вверх в следствии возмущения , то в соседнем слое архимедова сила станет больше силы тяжести , так как 2 1 . В верхней части малый объем , смещаясь вниз , поподает в облость пониженной плотности , и архимедова сила будет меньше силы тяжести FA < FT , возникает нисходящее движение жидкости . Направление движения нисходящего и восходящего потоков в данной ячейке случайно , движение же потоков в соседних ячейках , после выбора направлений в данной ячейке детерминировано . Полный поток энтропии через границы системы отрицателен , то есть система отдает энтропию , причем в стационарном состоянии отдает столько , сколько энтропии производится внутри системы (за счет потерь на трение). dSe q q T1 - T2 = - = q < 0 (3.1) dt T2 T1 T1 T2 Образование именно сотовой ячеистой структуры объясняется минимальными затратами энергии в системе на создание именно такой формы пространственной структуры . При этом в центральной части ячейки жидкость движется вверх , а на ее периферии - вниз. Дальнейшее сверхкритическое нагревание жидкости приводит к разрушению пространственной структуры - возникает хаотический турбулентный режим.  Рис. 3.2. Иллюстрация возникновения тепловой конвекции в жидкости . К этому вопросу прикладывается наглядная иллюстрация возникновения тепловой конвекции в жидкости . ЛАЗЕР , КАК САМООРГАНИЗУЮЩАЯСЯ СИСТЕМА. Во второй главе этот вопрос мы уже рассматривали . Здесь же , рассмотрим простую модель лазера . Лазер - это устройство , в котором в процессе стимулированного излучения порождаются фотоны . Изменение со временем числа фотонов n , или другими словами , скорость порождения фотонов , определяется уравнением вида : dn / dt = «Прирост» - «Потери» (3.2) Прирост обусловлен так называемым стимулированном излучением . Он пропорционален числу уже имеющихся фотонов и числу возбужденных атомов N . Таким образом : Прирост = G N n (3.3) Здесь G - коэффициент усиления , который может быть получен из микроскопической теории . Член , описывающий потери , обусловлен уходом фотонов через торцы лазера . Единственное допущение , которое мы принимаем , - это то , что скорость ухода пропорциональна числу имеющихся фотонов . Следовательно , Потери = 2n (3.4) 2 = 1/ t0 , где t0 - время жизни фотона в лазере . Теперь следует учесть одно важное обстоятельство , которое делает (2.1) нелинейным уравнением вида : (3.5) Число возбужденных атомов уменьшается за счет испускания фотонов . Это уменьшение N пропорционально числу имеющихся в лазере фотонов , поскольку эти фотоны постоянно заставляют атомы возвращаться в основное состояние . N = n (3.6) Таким образом , число возбужденных атомов равно N = N0 - N (3.7) где N0 - число возбужденных атомов , поддерживаемое внешней накачкой , в отсутствии лазерной генерации. Подставляя (3.3) - (3.7) в (3.2) , получаем основное уравнение нашей упрощенной лазерной модели : (3.8) где постоянная k дает выражение : k1 = G k = 2 - GN0 0 (3.9) Если число возбужденных атомов N0 (создаваемых накачкой) невелико , то k положительно , в то время как при достаточно больших N0 k - может стать отрицательным . Изменение знака происходит когда GN0 = 2 (3.10) Это условие есть условие порога лазерной генерации . Из теории бифуркации следует , что при k > 0 лазерной генерации нет , в то время как при k < 0 лазер испускает фотоны. Ниже или выше порога лазер работает в совершено разных режимах . Решим уравнение (3.8) и проанализируем его аналитически : - это уравнение одномодового лазера . Запишем уравнение (3.8) в следующем виде : Разделим исходное уравнение на n2 . и введем новую функцию Z : 1/n = n-1 = Z Z1 = - n-2 следовательно уравнение примет вид : перепишем его в следующем виде : разделим обе части данного уравнения на -1 , получим (3.11) Уравнение (3.11) - это уравнение Бернулли , поэтому сделаем следующую замену Z = UV , где U и V неизвестные пока функции n , тогда Z1 = U1 V + U V1 . Уравнение (3.11) , после замены переменных , принимает вид U1 V + UV1 - k UV = k1 преобразуем , получим U1 V + U(V1 - k V) = k1 (3.12) Решим уравнение (3.12) V1 - k V = 0 dV/dt = k V сделаем разделение переменных dV/V =k dt ln V = k t результат V = ekt (3.13) Отсюда мы можем уравнение (3.12) переписать в виде : U1 ekt = k1 - это то же самое , что dU/dt = k1e-kt , dU = k1e -kt dt выразим отсюда U , получим (3.14) По уравнению Бернулли мы делали замену Z = U V подставляя уравнения (3.13) и (3.14) в эту замену , получим Ранее вводили функцию Z = n-1 , следовательно (3.15) Начальное условие n0=1/(c-k1/k) , из этого условия мы можем определить константу с следующим образом Подставляя , найденную нами константу в уравнение (3.15) , получим (3.16) Исследуем функцию (3.16) при k = 0 , k < 0 , k > 0 . При k0 ; ekt 0 ; (ekt - 1)0 , то есть (ekt - 1)k1/k0 (неопределенность) , раскроем эту неопределенность по правилу Лопиталя . Эту неопределенность вида 0 следует привести к виду . При этом , как и всегда при применении правила Лопиталя , по ходу вычислений рекомендуется упрощать получившиеся выражения , следующим образом : n(k)при k0 0 , следовательно Перепишем (3.16) в следующем виде Линеаризуем нелинейное уравнение , получим ln n = - kt + c Построим график для этих условий 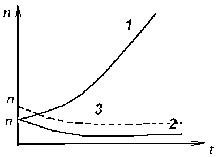 Рис. 3.3 К самоорганизации в одномодовом лазере : кривая 1 : k < 0 , режим лазерной генерации кривая 2 : k = 0 , точка бифуркации , порог кривая 3 : k > 0 , режим лампы. При k = 0 уравнение (3.8) примет вид решая его , получим (3.8) При условии ; n(t) = const , функция (3.8) приближается к стационарному состоянию , не зависимо от начального значения n0 , но в зависимости от знаков k и k1 (смотри рисунок 3.3). Таким образом , функция (3.8) принимает стационарное решение 3.3. ДИНАМИКА ПОПУЛЯЦИИ . О распространении и численности видов была собрана обширная информация . Макроскопической характеристикой , описывающей популяцию , может быть число особей в популяции . Это число играет роль параметра порядка . Если различные виды поддерживаются общим пищевым ресурсом , то начинается межвидовая борьба , и тогда применим принцип Дарвина : выживает наиболее приспособленный вид . ( Нельзя не отметить сильнейшую аналогию , существующую между конкуренцией лазерных мод и межвидовой борьбой ). Если имеются однотипные пищевые ресурсы , то становится возможным сосуществование видов . Численность видов может быть подвержена временным колебаниям. ОДИН ВИД. Рассмотрим сначала одну популяцию с числом особей в ней n . При наличии пищевых ресурсов А особи размножаются со скоростью : и гибнут со скоростью : Здесь k и d - некоторые коэффициенты рождаемости и смертности , в общем случае зависящее от параметров внешней среды обитания . Если бы количество пищи было неограниченно , то эволюционное уравнение выглядело бы так : Введем обозначение = kA - d Оно было бы линейным и описывало бы неограниченный экспериментальный рост (при kA > d), либо экспериментальную гибель (при kA < d) популяции. 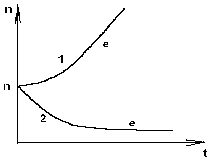 Рис. 3.4 Кривая 1: Экспоненциальный рост ; >0 , kA>d Кривая 2: Экспоненциальная гибель ; >0 , kA>d. В общем случае , однако , пищевые ресурсы ограничены , так что скорость потребления пищи Вместе с тем в общем случае возможно восстановление пищевых ресурсов со скоростью : Здесь , конечно , рассмотрен придельный случай сохранения полного количества органического вещества A + n = N = const , N - способность среды обитания поддерживать популяцию. Тогда с учетом A = N - n получится следующее уравнение эволюции популяции одного вида (логистическое уравнение Ферхюльста ) : (3.17) Решим уравнение (3.17) аналитически , перепишем его следующим образом , обозначим kN - d = k1 Получим : Воспользуемся табличным интегралом , ,полученное уравнение примет вид : решим это уравнение , преобразуя сократим полученное выражение на k , и перенесем переменную k1 в правую часть , получим отсюда n(t) Начальные условия : откуда Подставляя с в решение , получим уравнение в следующем виде ранее мы обозначали , что , подставляем и преобразуем сократим на k - коэффициент рождаемости , окончательно получим решение уравнения (3.17) Итак , получено аналитическое решение логистического уравнения - это решение указывает на то , что рост популяции останавливается на некотором конечном стационарном уровне: то есть параметр n1 указывает высоту плато насыщения , к которому стремится n(t) с течением времени . Параметр n0 указывает начальное значение численности одного вида популяции : n0 = n(t0) . Действительно , ,то есть n1 - предельная численность вида в данной среде обитания . Иначе говоря , параметр n1 характеризует емкость среды по отношению к данной популяции . И наконец , параметр (kN - d) задает крутизну начального роста . Отметим , что при малой исходной численности n0 (начальное число особи) начальный рост популяций будет почти экспоненциальным 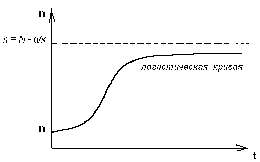 Рис. 3.5. Логистическая кривая. (эволюция популяции одного вида) Решение уравнения (3.17) можно представить с помощью логистической кривой (рис. 3.5) . Эволюция полностью детерминирована . Популяция перестает расти , когда ресурс среды оказывается исчерпанным . Самоорганизация - при ограниченном пищевом ресурсе. Система самоорганизованна и взрывоподобный рост популяции (рис. 3.4 Кривая 1) сменяется кривой с насыщением . Подчеркнем , что при описании данной биологической системы используют понятийный и физико-математический аппарат из нелинейной неравновесной термодинамики. Может случится , однако, что всегда за событиями , не управляемыми в рамках модели , в той же среде появится , первоначально в малых количествах , новые виды (характеризуемые другими экологическими параметрами k,N и d) . В связи с такой экологической флуктуацией возникает вопрос о структурной устойчивости : новые виды могут либо исчезнуть , либо вытеснить первоначальных обитателей . Пользуясь линейным анализом устойчивости , не трудно показать , что новые виды вытесняют старые только в том случае , если Последовательность , в которой виды заполняют экологическую нишу , представлена на рисунке 3.6. 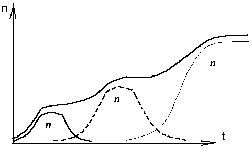 Рис. 3.6. Последовательное заполнение экологической ниши различными видами . Эта модель позволяет придать точным количественный смысл утверждению о том , что «выживает наиболее приспособленный» , в рамках задачи о заполнении заданной экологической ниши . СИСТЕМА «ЖЕРТВА - ХИЩНИК». Рассмотрим систему, состоящую из двух видов - это «жертва» и «хищник» (например , зайцы и лисицы) , то эволюция системы и ее самоорганизация выглядят иначе , чем в предыдущем случае. Пусть в биологической системе имеются две популяции - «жертв» - кролики (К) , и «хищников» - лисиц (Л), численностью К и Л . Проведем теперь рассуждение , которое позволит нам объяснить существование диссипативных структур . Кролики (К) поедают траву (Т) . Предположим , что запас травы постоянен и неисчерпаем . Тогда , одновременное наличие травы и кроликов способствуют неограниченному росту кроличьей популяции . Этот процесс можно символически изобразить так : Кролики + Трава Больше кроликов К + Т 2К Тот факт , что в стране кроликов всегда имеется в достатке травы , вполне аналогичен непрерывному подводу тепловой энергии в задаче с ячейками Бенара . Вскоре процесс , в целом , будет выглядеть как диссипативный (во многом аналогично процессу Бенара ). Реакция « Кролики - Трава » происходит спонтанно в направлении увеличения популяции кроликов, что является прямым следствием второго начала термодинамики . Но вот в нашу картину , где мирно резвятся кролики , прокрались хищные лисицы (Л), для которых кролики являются добычей . Подобно тому , как по мере поедания травы кроликов становится больше , за счет поедания кроликов возрастает число лисиц : Лисицы + Кролики Больше лисиц Л + К 2Л В свою очередь лисицы , как и кролики являются жертвами - на этот раз человека , точнее говоря происходит процесс Лисицы Меха Конечный продукт - Меха , не играет непосредственной роли в дальнейшем ходе процесса . Этот конечный продукт можно , однако , рассматривать как носитель энергии, выводимой из системы , к которой она была в начале подведена (например, в виде травы ). Таким образом , в экологической системе также существует поток энергии - аналогично тому , как это имеет место в химической пробирке или биологической клетке . Совершенно ясно , что в действительности происходят периодические колебания численности популяции кроликов и лисиц , причем за нарастании численности кроликов следует нарастание численности лисиц , которые сменяются уменьшением численности кроликов , сопровождающимся столь же резким снижением численности лисиц , затем повышенным подъемом численности кроликов и так далее (рис. 3.7). 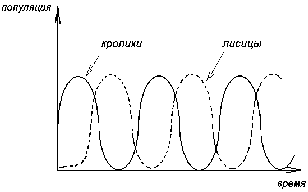 Рис. 3.7. Изменение численности популяций кроликов и лисиц со временем. Наличие периодичности означает возникновение экологической структуры. С течением времени численность обеих популяций меняется в соответствии с последовательным прохождением точек графика . Через некоторое время (конкретное значение зависит от быстроты поедания лисицами кроликов , а так же от скорости размножения обоих видов) весь цикл начинается вновь. Поведение популяций при различных степенях плодовитости , а так же различных способностях избегать истребления можно изучить количественно с помощью программы : ПОПУЛЯЦИЯ (в приложении). Эта программа реализует решение уравнений для диссипативной структуры «кролики - лисицы». Результат решения изображается графически . Решается система дифференциальных уравнений Здесь буквы К, Л, Т - означают соответственно количество кроликов , лисиц , травы ; коэффициенты k1, k2, k3 - обозначают соответственно скорость рождения кроликов , скорость поедания кроликов лисицами и скорость гибели лисиц. В программе понадобится уточнить значение отношений |