Физиология. 3 пара по физе. 1. Анатомогистологическая и функциональная классификация нервных волокон
 Скачать 0.94 Mb. Скачать 0.94 Mb.
|

|
1.Анатомо-гистологическая и функциональная классификация нервных волокон Нервные волокна — это отростки нейронов, с помощью которых осуществляется связь между нейронами и с исполнительными клетками. Имеется два типа нервных волокон: миелинизированные и немиелинизированные (безмиелиновые). Оболочку безмиелиновых волокон образуют шванновские клетки (леммоциты), в которые погружаются осевые цилиндры нервных волокон. Оболочку миелинизированных волокон образуют в периферической нервной системе также шванновские клетки (миелоциты, глиальная клетка), формирующие миелин (многослойная обертка мембран миелоцитов — до 100), а в ЦНС — олигодендроциты (глиальные клетки, т.е. те же шванновские клетки).
Миелиновая оболочка через равные участки (около 1 мм) прерывается, образуя свободные от миелина небольшие участки — перехваты Ранвье. Основную часть миелина (78% сухого веса) составляют липиды, обеспечивающие изолирующие свойства оболочки. Нервные волокна обеспечивают проведение возбуждения и аксонный транспорт, выполняющий трофическую функцию нейрона. *Миелинизация нервных окончаний - это образование специального слоя вещества под названием "миелин" вокруг осевого цилиндра нервного волокна в период его созревания. 2. Особенности проведения возбуждения по миелиновым и безмиелиновым волокнам Механизм распространения возбуждения у различных нервных волокон неодинаков. Механизмы проведения возбуждения в безмиелиновых волокнах. При действии раздражителя пороговой силы на мембрану безмиелинового волокна изменяется ее проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны (внутренняя становиться заряженной положительно, а наружная отрицательно). Это ведет к возникновению круговых токов (заряженных частиц) от «+» к «–» на протяжении всего волокна. Особенности распространения возбуждения по безмиелиновым волокнам: 1. Возбуждение распространяется непрерывно и все волокно сразу охватывается возбуждением. 2. Возбуждение распространяется с небольшой скоростью. 3. Возбуждение распространяется с декриментом (уменьшение силы тока к концу нервного волокна). По безмиелиновым волокнам возбуждение проводится к внутренним органам от нервных центров. Однако низкая скорость распространения возбуждения и его затухание не всегда выгодно организму. Поэтому природой был выработан еще один дополнительный механизм распространения возбуждения. 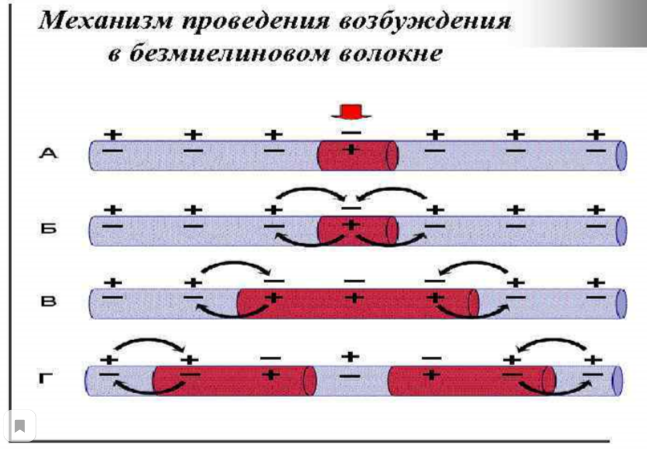 Механизмы проведения возбуждения в миелиновых волокнах. Наличие у миелиновых волокон оболочки, обладающей высоким электрическим сопротивлением, а также участков волокна, лишенных оболочки - перехватов Ранвье создают условия для качественно нового типа проведения возбуждения по миелиновым нервным волокнам. В миелинизированномволокне токи проводятся только в зонах, не покрытых миелином (перехватах Ранвье). В этих участках генерируется очередной ПД. Перехваты длиной 1 мкм расположены через 1000 - 2000 мкм, характеризуются высокой плотностью ионных каналов, высокой электропроводностью и низким сопротивлением. При действии раздражителя пороговой силы на мембрану миелинового волокна в области перехвата Ранвье изменяется проницаемость для ионов Nа+, которые мощным потоком устремляются внутрь волокна. В этом месте изменяется заряд мембраны, что ведет к возникновению круговых токов. Этот ток идет через межтканевую жидкость к соседнему перехвату, где происходит смена заряда. Таким образом, возбуждение перепрыгивает с одного участка на другой. Обратное движение возбуждения невозможно так как участок, через который оно прошло, находится в фазе абсолютной рефрактерности. Особенности распространения возбуждения по миелиновым волокнам: 1. Распространение ПД в миэлинизированных нервных волокнах осуществляется сальтаторно - скачкообразно от перехвата к перехвату, т.е. возбуждение (ПД) как бы «перепрыгивает» через участки нервного волокна, покрытые миелином, от одного перехвата к другому и все волокно сразу не охватывается возбуждением. 2. Возбуждение распространяется с большой скоростью. 3. Возбуждение распространяется без декримента. По миелиновым волокнам возбуждение распространяется от анализаторов к ЦНС, к скелетным мышцам, т.е. там, где требуется высокая скорость ответной реакции. 3.Законы проведения возбуждения по нервным волокнам Первый закон – «Закон анатомической и физиологической непрерывности» Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву. Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т.д.) приводят к нарушению физиологической целостности. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается. Второй закон – «Закон двустороннего проведения возбуждения по нерву» От места возникновения, возбуждение может распространяться по нерву в обе стороны от места раздражения. В естественных условиях возбуждение распространяется по афферентным проводникам – к клетке, а по эфферентным – от клетки. Оно носит название ортодромного проведения. Обратное направление движения возбуждения, редко наблюдаемое в организме (например при антидромной тахикардии), называют антидромным проведением. Третий закон – «Закон изолированного проведения возбуждения по нервному волокну». В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервного волокна на другое, то нормальное функционирование органов было бы невозможно. В учебнике есть еще закономерности, но в инете их нет так тож хз смотрите сами: 4) Возбуждение распространяется бездекрементно (не затухая). Это обусловлено тем, что локальные токи лишь деполяризуют мембрану до критического уровня, а потенциал действия возникает регенеративно за счет трансмембранных ионных перемещений, перпендикулярных к направлению проведения самого возбуждения. 5) Скорость проведения возбуждения тем больше, чем выше амплитуда потенциала действия. Т.к. при этом возрастает разность потенциалов возбужденного и невозбужденного участков мембраны. 6) Скорость проведения возбуждения прямопропорциональна диаметру нервного волокна. Т.к. с увеличением диаметра уменьшается сопротивление. 4.Строние нервно-мышечного синапса. Формирование потенциала концевой пластинки. Отличия механизма возникновении потенциала концевой пластинки от механизма формирования потенциала действия. Химическая передача возбуждения между возбудимыми клетками является основным способом и реализуется с помощью синапсов. Синапс - это место контакта между мембраной терминали нервной клетки, с одной стороны, и мембраной другой возбудимой клетки (нейрон, мышечная клетка), которое обеспечивает передачу потенциалов действия от одной клетки к другой с помощью молекул химических веществ, или медиаторов. Передача возбуждения с нерва на мышцу осуществляется через нервно-мышечный синапс, который представляет собой место контакта окончания аксона двигательного нейрона с мембраной мышечного волокна. Нервно-мышечный синапс состоит из пресинаптической и постсинаптической мембраны, между которыми имеется пространство в 10-20 нм, которое называется синаптической щелью. Пресинаптическая мембрана нервно-мышечного синапса представляет собой мембрану терминали аксона мотонейрона спинного мозга, а постсинаптическая мембрана является мембраной мышечного волокна. Постсинаптическая мембрана нервно-мышечного синапса называется также двигательной концевой пластинкой. Передача возбуждения через нервно-мышечный синапс происходит в несколько условно разделенных этапов, включающих пресинаптический и постсинаптический механизмы передачи возбуждения. Постсинаптический механизм передачи возбуждения через нервно-мышечный синапс трансформирует химический сигнал (количество молекул ацетилхолина) в электрический сигнал. Молекулы ацетилхолина кратковременно (не более 2 мс) взаимодействуют с N-холинорецепторами постсинаптической мембраны, открывая натриевые и калиевые ионные каналы в каждом рецепторном белке. Плотность N-холинорецепторов на постсинаптической мембране выше, чем в синапсах нервной системы (1х104 на 1 мкм2), что обеспечивает высокую надежность функции передачи возбуждения через нервно-мышечный синапс. Ионы Na по электрохимическому градиенту диффундируют из синаптической щели внутрь мышечного волокна, а ионы К - в обратном направлении. Однако входящий ток ионов натрия преобладает над выходящим калиевым током. В результате происходит деполяризация постсинаптической мембраны нервно-мышечного синапса, которая называется потенциалом концевой пластинки. Продолжительность этого потенциала очень мала - не более 1-2 мс, а его амплитуда варьирует в зависимости от количества высвобождающегося ацетилхолина. При этом между амплитудой потенциала концевой пластинки и количеством высвобождающегося ацетилхолина имеется прямая зависимость. Величины одного потенциала концевой пластинки достаточно для возбуждения мембраны мышечного волокна вблизи мембраны двигательной концевой пластинки. Амплитуда потенциала концевой пластинки, достигая по величине порога возбуждения мышечной мембраны (-15...-20 мВ), вызывает генерацию в ней потенциала действия, который распространяется по мембране мышечного волокна. Поскольку большинство двигательных концевых пластинок расположено в средней части мышечного волокна, то потенциал действия распространяется по его мембране в двух направлениях, т. е. к концам волокна. Как только потенциал действия, распространяясь по сарколемме мышечного волокна, достигает его t-системы, ионы Са начинают высвобождаться из саркоплазматического ретикулума, и в мышце происходит сокращение. В зависимости от количества генерируемых в мышечном волокне потенциалов действия сокращение может быть одиночным или суммированным. Надежность передачи возбуждения в нервно-мышечном синапсе настолько высока, что каждый потенциал действия двигательного нейрона спинного мозга вызывает генерацию одного потенциала действия в каждом мышечном волокне двигательной единицы. После прекращения активности двигательного нейрона спинного мозга ионы Са с помощью Са-АТФазы и Na-, Са-обменника выводятся из цитоплазмы нервного окончания во внеклеточную среду. В нервном окончании везикулы заполняются вновь синтезированными молекулами ацетилхолина. На постсинаптической мембране, благодаря работе Na-, K-АТФаз, восстанавливаются ионные градиенты Na и K, а также нормальная возбудимость постсинаптической мембраны. 1.Морфо-функциональные характеристики скелетной мышцы: саркомер, саркоплазматический ретикулум, сократительные и регуляторные белки. Саркомер — базовая сократительная единица поперечнополосатых мышц, представляющая собой комплекс нескольких белков, состоящий из трёх разных систем волокон. Из саркомеров состоят миофибриллы. Саркомер - это участок миофибриллы между двумя соседними Z-линиями, структурно-функциональная единица поперечнополосатой мышечной ткани и других ее составляющих компонентов 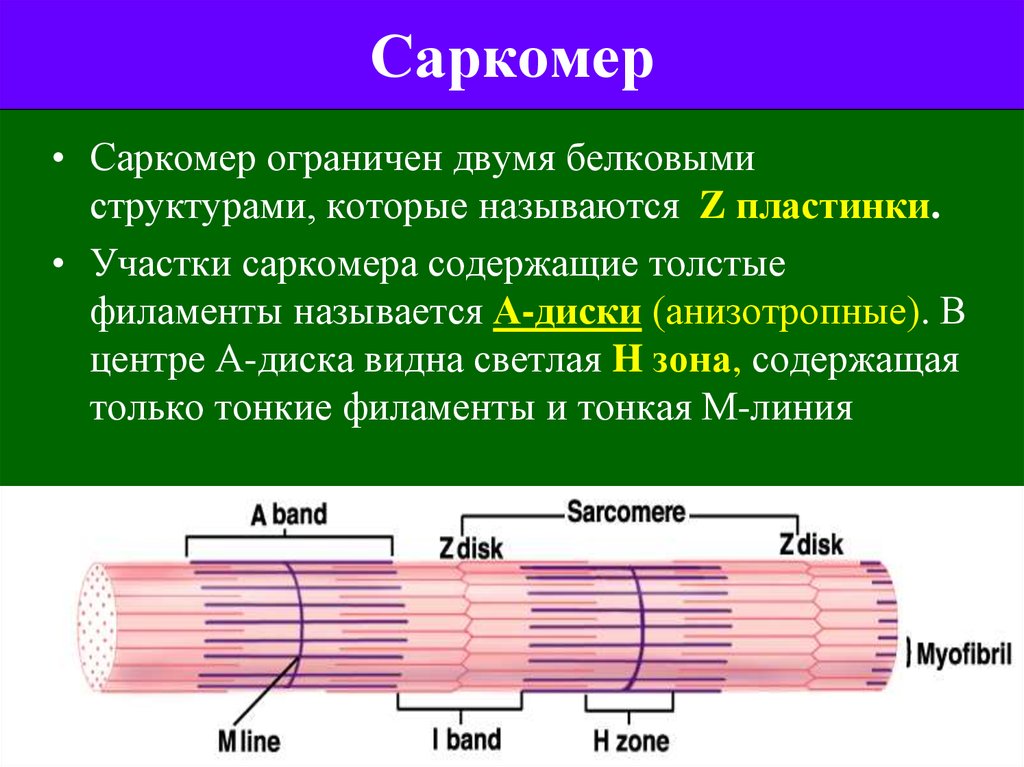 Саркоплазматический ретикулум (гладкая эндоплазматическая сеть) – мембранная органелла, представляющая собой систему трубочек и мешочков (цистерн), окружающих миофибриллы. Известно, что миофибриллы не имеют оболочки, поэтому функцию оболочки выполняет саркоплазматический ретикулум, который окружает каждую миофибриллу наподобие «муфточки» или «кружевного рукава» 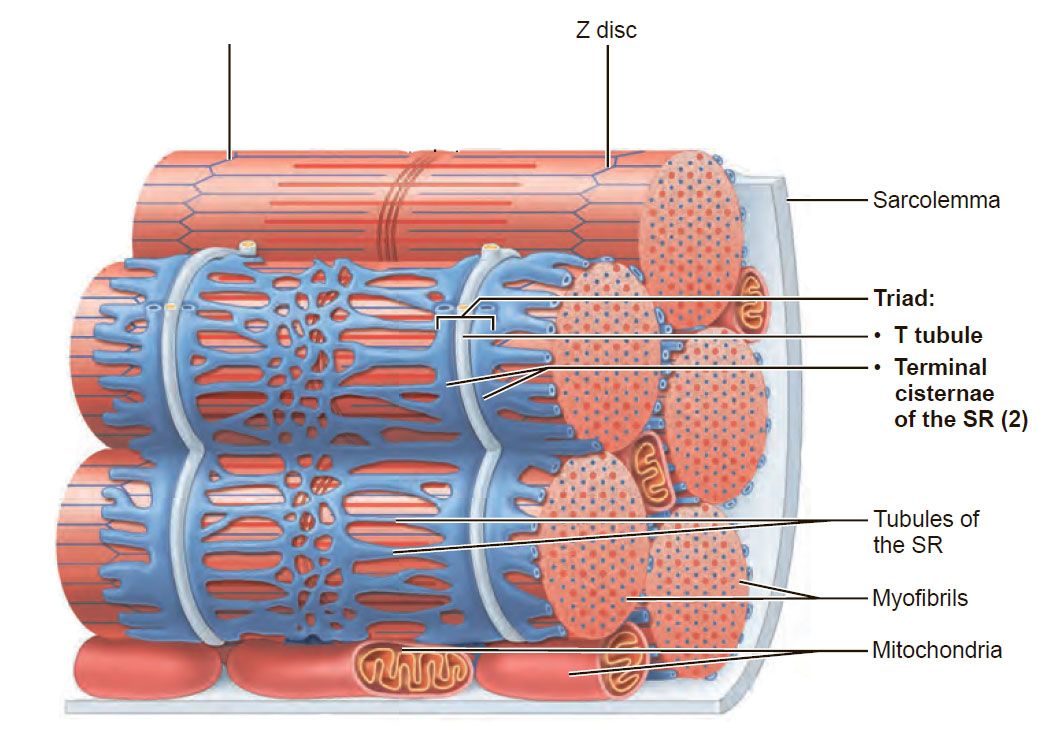 Структура Саркоплазматический ретикулум представляет собой единую систему компонентов различной формы — от трубочек до уплощенных цистерн. Благодаря ответвлениям продольные каналы, окружающие каждую миофибриллу, соединяются друг с другом, а также с другими каналами, окружающими другие миофибриллы. Функция Основная функция саркоплазматического ретикулума – депонирование и выделение ионов кальция (Са2+). В состоянии покоя в саркоплазматическом ретикулуме депонируются ионы кальция. В саркоплазме мышечного волокна концентрация этих ионов очень низкая. В начале сокращения мышечного волокна продольные каналы саркоплазматического ретикулума становятся шире и короче и из него в саркоплазму выделяются ионы кальция, необходимых для процесса сокращения мышечного волокна. После окончания процесса сокращения мышечного волокна ионы кальция закачиваются в саркоплазматический ретикулум посредством кальциевого насоса. Т-система От поверхности мышечного волокна к расширенным участкам саркоплазматического ретикулума направляются выпячивания сарколеммы – поперечные трубочки, называемые Т-системой (рис.2). Главная функция трубочек – проведение возбуждающих импульсов с поверхности мышечного волокна в его центральную зону, что приводит к выделению ионов кальция из саркоплазматического ретикулума. 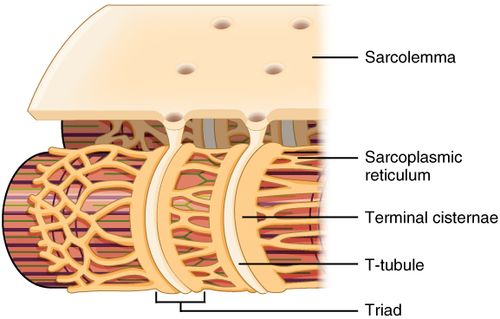 Эндоплазматическая сеть мышечного волокна Повреждение мышечных волокон При повреждении мышечных волокон очень часто повреждается саркоплазматический ретикулум. Это приводит к значительному выбросу ионов кальция в саркоплазму мышечного волокна и активации ферментов – протеаз, разрушающих белки. 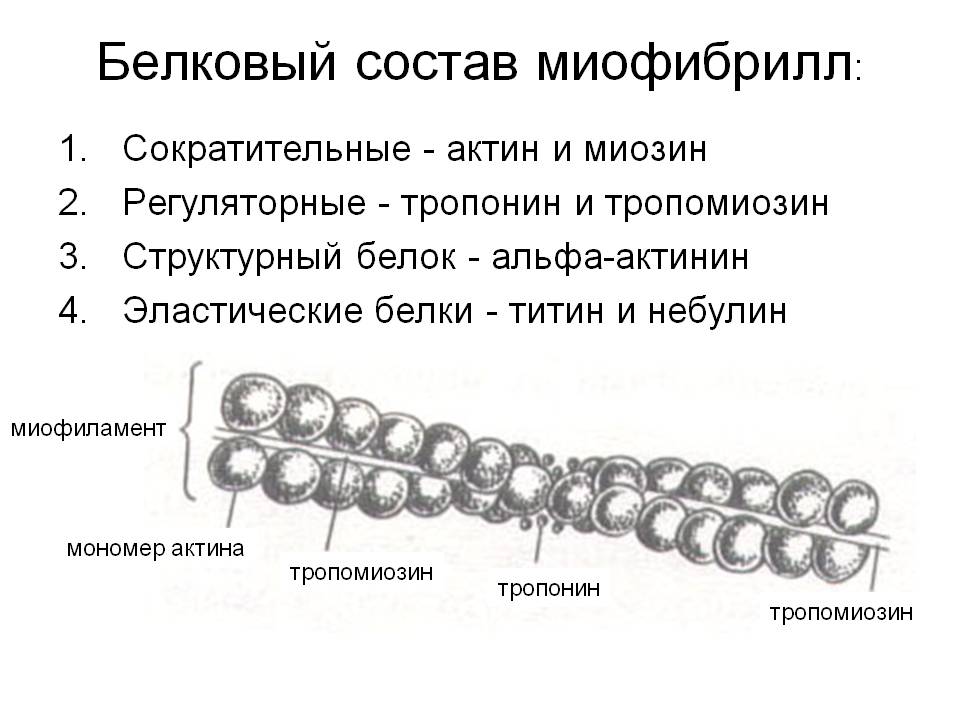 Актин — глобулярный белок, из которого образованы микрофиламенты — один из основных компонентов цитоскелета эукариотических клеток. Вместе с белком миозином образует основные сократительные элементы мышц — актомиозиновые комплексы саркомеров. Присутствует в основном в цитоплазме, но в небольшом количестве также найден в ядре клетки. Функции - Формируют клеточный цитоскелет, создавая механическую поддержку. - Принимает участие в миозин-независимом изменении формы клетки и клеточное движение. - В мышечных клетках актин вместе с миозином создает комплекс, участвующий в сокращении мышцы. - В немышечных клетках принимает участие в транспортировке миозином везикул и органелл - Деление клеток и цитокинез Тропонин — регуляторный глобулярный белок, состоящий из трех субъединиц, который участвует в процессе мышечного сокращения. Содержится в скелетных мышцах и сердечной мышце, но не содержится в гладкой мускулатуре. В ряде медицинских тестов тропонин используется как биомаркер различных сердечных заболеваний. Тропомиозин — фибриллярный белок. Тропомиозин совместно с тропонином участвует в регуляции взаимодействия актина с миозином. №2. Механизм электромеханического сопряжения, теория скольжения: роль ионов кальция, регуляторных и сократительных белков, головок миозина и АТФ. Электромеханическое сопряжение – это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску сокращения мышцы. Кроме сократительных белков актина и миозина, в мышцах присутствуют регуляторные белки тропонин и тропомиозин. В состоянии покоя они закрывают активные центры молекулы актина, и из-за этого взаимодействие актиновых и миозиновых нитей в состоянии покоя не происходит. Когда ионы Са доходят до межфибрилярного пространства, они взаимодействуют с регуляторными белками и изменяют конформацию регуляторных белков, открываются актиновые центры и миозин быстро вступает в реакцию, что ведет к образованию акто-миозинового комплекса. Т.е головки миозина становятся практически перпендикулярными и совершают гребковое движение. Энергия от АТФ будет расходоваться на расщепление этого акто-миозинового комплекса с образованием вновь свободного актинового центра и белка миозина. Но головка миозина уже будет расположена у следующего актина и опять произойдет передвижение. Этот механизм движения актиновых и миозиновых нитей и называют механизмом скольжения. 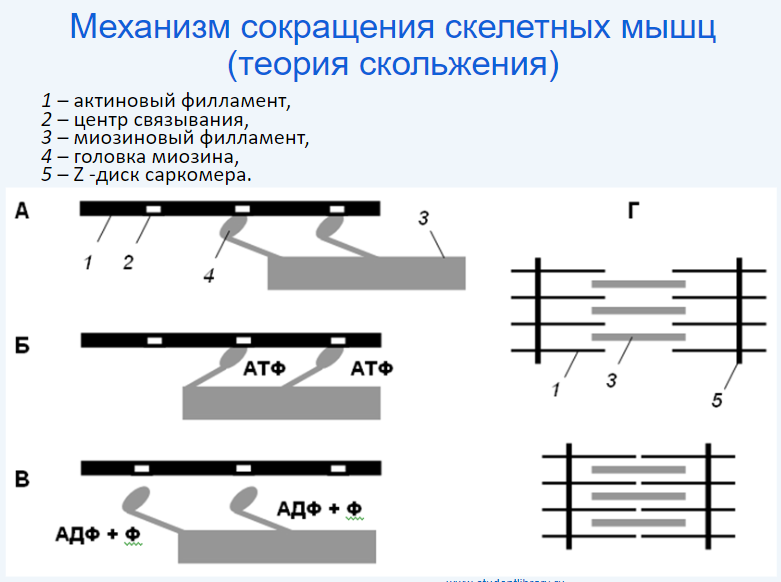  3.Режимы сокращения изолированных скелетных мышц: одиночное сокращение зубчатый и гладкий тетанус. Раздражение мышцы одиночным пороговым стимулом приводит к возникновению одиночного сокращения , которое состоит из трёх периодов: 1) латентного периода; 2) периода укорочения); 3) периода расслабления. 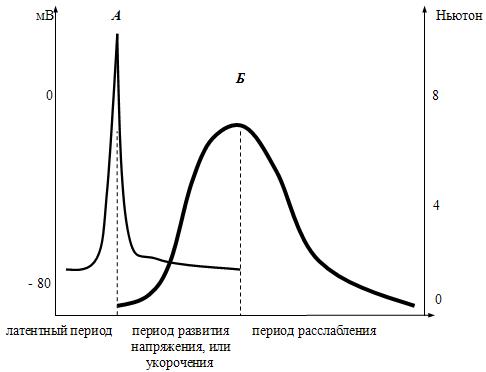 Рис. 10. Периоды одиночного мышечного сокращения: А – потенциал действия; Б – мышечное сокращение. 1) латентный период– это время от момента нанесения раздражителя до начала укорочения МВ и обусловлен возбуждением мембраны мышечного волокна, повышением концентрации внутриклеточного кальция и активации поперечных мостиков. Он составляет около 2 мс (0,02с). 2)период укорочения или развития напряжения. Он длится 0,05 мс. В случае свободного укорочения мышечного волокна говорят об изотоническом режиме сокращения, при котором меняется только длина мышечного волокна, а напряжение остается относительно постоянным. Если мышечное волокно закреплено с двух сторон и не может свободно укорачиваться, то говорят об изометрическом режиме сокращения. В этом случае МВ укоротиться больше не может, а возникающее напряжение передается на эластические элементы (поперечные мостики миозиновых нитей, актиновые нити, Z-пластинки, продольно расположенная саркоплазматическая сеть и сарколемма мышечного волокна), расположенные внутри волокна. В процессе жизнедеятельности в организме человека развивается комбинация изотонического и изометрического компонентов - ауксотонический режим сокращения. Как правило, укорочение мышцы сопровождается развитием напряжения и увеличением силы, достаточной чтобы совершить перемещение груза. 3) период расслабления (0,07 мс), когда уменьшается концентрация ионов Са2+ и отсоединяются головки миозина от актиновых филаментов. Если нанести на скелетную мышцу дополнительно второй, третий и т.д. раздражающие стимулы в период расслабления, то происходит суммирование возникающих сокращений и результирующее сокращение по силе будет выше, чем сокращение на одиночный стимул – это неполная суммация одиночных сокращений. Картина этих сокращений называется зубчатым тетанусом , т.к. на кривой записи видны характерные западения (рис.11А). Тетанус – это сильное и длительное сокращение мышцы. У здорового человека сокращения мышц по типу зубчатого тетануса не встречаются. 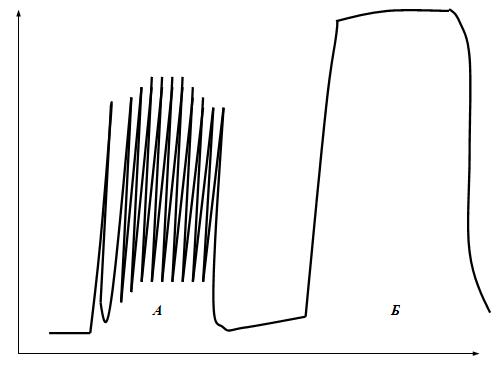 Рис. 11. Зубчатый (А) и гладкий (Б) тетанусы. При увеличении частоты дополнительные стимулы приходятся на период укорочения (или напряжения), и происходит полная суммация одиночных сокращений, которая называется гладким тетанусом (рис.11Б). На кривой записи такого мышечного сокращения нет западений, она гладкая. Амплитуда этой кривой выше, чем у кривых одиночного сокращения и зубчатого тетануса. При этом чем больше частота раздражения (в определённых пределах), тем выше амплитуда тетанического сокращения. Та частота, при которой амплитуда тетануса максимальная, называется оптимальной. Частоты, которые больше оптимальной, называются пессимальными, амплитуда тетануса при этом снижается. В организме к скелетным мышцам в естественных условиях всегда приходит серия импульсов, которые попадают в период укорочения мышцы, т.е. скелетные мышцы человека всегда сокращаются по типу гладкого тетануса. Существует две причины увеличения силы тетанических сокращений в естественных условиях: 1) увеличение числа возбуждённых мотонейронов и синхронизация частоты их возбуждений. Чем больше возбуждённых мотонейронов, тем больше сокращающихся двигательных единиц (это явление пространственной суммации сокращений моторных единиц). Чем больше степень синхронизации мотонейронов, тем больше амплитуда при суперпозиции максимального сокращения, развиваемого каждой двигательной единицы в отдельности; 2) увеличение частоты импульсов, генерируемых каждым мотонейроном (явление временной суммации сокращений каждого волокна данной моторной единицы). 4.Типы мышечного сокращения : Изотонический режим — сокращение, при котором мышца укорачивается без формирования напряжения. Такое сокращение возможно при пересечении или разрыве сухожилия или в эксперименте на изолированной (удаленной из организма) мышце. Изометрический режим — сокращение, при котором напряжение мышцы возрастает, а длина практически не уменьшается. Такое сокращение наблюдается при попытке поднять непосильный груз. Ауксотонический режим — сокращение, при котором длина мышцы изменяется по мере увеличения ее напряжения. Такой режим сокращений наблюдается при осуществлении трудовой деятельности человека. Если напряжение мышцы возрастает при ее укорочении, то такое сокращение называют концентрическим, а в случае увеличении напряжения мышцы при ее удлинении (например, при медленном опускании груза) — эксцентрическим сокращением. 5.Физиологические особенности и функции гладких мышц. По структуре гладкая мышца отличается от поперечнополосатой скелетной мышцы и мышцы сердца. Она состоит из клеток длиной от 10 до 500 мкм, шириной 5-10 мкм, содержащих одно ядро. Гладкие мышцы подразделяют, главным образом, на два типа: мультиунитарные и унитарные. Мультиунитарная гладкая мышца: состоит из отдельных гладкомышечных клеток, каждая из которых, находится независимо друг от друга; имеет большую плотность иннервации; как и поперечнополосатые мышечные волокна, снаружи покрыты веществом, напоминающим базальную мембрану, в состав которого входят, изолирующие клетки друг от друга, коллагеновые и гликопротеиновые волокна; каждая мышечная клетка может сокращаться отдельно и ее активность регулируется нервными импульсами. Унитарная гладкая мышца (висцеральная): представляет собой пласт или пучок, а сарколеммы отдельных миоцитов имеют множественные точки соприкосновения. Это позволяет возбуждению распространяться от одной клетки к другой; мембраны рядом расположенных клеток образуют множественные плотные контакты (gap junctions), через которые ионы имеют возможность свободно передвигаться из одной клетки в другую; потенциал действия, возникающий на мембране гладкомышечной клетки, и ионные потоки могут распространяться по мышечному волокну, обеспечивая возможность одновременного сокращения большого количества отдельных клеток. Данный тип взаимодействия известен как функциональный синцитий. Особенности электронномикроскопического строения гладкомышечных клеток В гладкой мышцы отсутствуют саркомеры. Толстые и тонкие миофиламеты распределены по всей саркоплазме гладкого миоцита и не имеют такой стройной организации, как в поперечно-полосатой скелетной мышце. При этом тонкие филаменты прикрепляются к плотным тельцам. Некоторые из этих телец расположены на внутренней поверхноти сарколеммы, но большинство из них находятся в саркоплазмме. Плотные тельца состоят из альфа-актинина – белка обнаруженного в структуре Z-мембраны поперечнополосатых мышечных волокон. Некоторые из плотных телец расположенных на внутренней поверхности мембраны соприкасаются с плотными тельцами прилегающей клетки. Тем самым сила, создаваемая одной клеткой может передаваться следующей. В гладкомышечных клетках практически отсутствуют Т-трубочки. Кроме того, гладкомышечные клетки значительно меньше поперечнополосатых мышечных волокон и поэтому не имеют развитой системы Т-трубочек, предназначенных для проведения возбуждения к расположенному в глубине сократительному аппарату. Вместо них наблюдаются небольшие углубления в сарколемме, которые получили название кавеолы. Благодаря им увеличивается площадь поверхности миоцита, а также может обеспечиваться взаимосвязь потенциалов, возникающих на мембране и саркоплазматическим ретикулумом. Особенности биопотенциалов гладкой мышцы Потенциал покоя. Величина разности потенциалов на мембране гладкомышечных клеток существенно различается в зависимоти как от типа гладкой мышцы так и от тех условий, в которых она находится. Обычно в состоянии покоя мембранный потенциал гладкомышечной клетки нестабилен и находится в пределах от –30 до –50 мВ, что на 30 мВ меньше, чем в скелетной мышце. Потенциал действия. Необходимо отметить, что в мембране гладкомышечной клетки обнаружено гораздо большее количество потенциалзависимых кальциевых каналов, чем в мембране поперечнополосатых мышечных волокон. Более того, ионы натрия играют малую роль в гененерации потенциала действия. Вместо них большое значение в генерации потенциала действия принадлежит потоку ионов кальция внутрь гладкомышечной клетки. Однако кальциевые каналы открываются значительно медленее, чем натриевые каналы, но остаются открытыми значительно дольше. На основании этого можно понять почему потенциал действия гладкой мышцы развивается в течение столь длительного времени. Другой важной задачей входящего во время потенциала действия кальция является их прямое влияние на сократительный аппарат клетки. Некоторые гладкомышечные клетки обладают способностью к самовозбуждению, то есть способны генерировать потенциал действия без воздействия внешнего раздражителя. Это часто связано с периодическими колебаниями мембранного потенциала. Очень часто подобная активность наблюдается в гладкой мышце кишечника. Медленные волновые колебания мембранного потенциала не являются потенциалом действия. Одним из возможных механизмов, объясняющих появление этих волновых колебаний мембранного потенциала, является периодическая активация и затухание активности натрий-калиевого насоса. Разность потенциалов на мембране гладкомышечной клетки увеличивается во время активации Na/K насоса и уменьшается при ее снижении. Другой возможной причиной данного явления является ритмическое увеличение или снижение проводимости ионных каналов. Физиологическое значение медленных колебаний мембранного потенциала состоит в том, что они могут инициировать появление потенциала действия. Это возникает в том момент когда во время медленной волны разность потенциалов на мембране клетки снижается до –35 мВ. При этом, как правило, успевает возникнуть несколько потенциалов действия. Следовательно, медленные волны можно назвать пейсмекерными волнами и, таким образом, становится понятным каким образом они обуславливают ритмические сокращения кишки. Одним из важных раздражителей, инициирующих сокращение гладких мышц, является их растяжение. Достаточное растяжение гладкой мышцы обычно сопровождается появлением потенциалов действия. Таким образом, появлению потенциалов действия при растяжении гладкой мышцы способствует два фактора: (1) медленные волновые колебания мембранного потенциала, на которые наслаивается (2) деполяризация, вызываемая растяжением гладкой мышцы. Данное свойство гладкой мышцы позволяет ей автоматически сокращаться при растяжении. Например, во время переполнения тонкого кишечника возникает перистальттическая волна, которая и продвигает содержимое. Деполяризация мультиунитарной гладкой мышцы. В обычных условиях мультиунитарная гладкая мышца сокращается в ответ на поступление нервного импульса. Чаще всего из нервного окончания высвобождается ацетилхолин, в некоторых мультиунитарных мышцах, норадреналин или другой медиатор. В любом случае медиатор приводит к деполяризации мембраны гладкой мышцы и к последующему ее сокращению. Потенциал действия при этом не возникает. Причина данного явления в том, что мультиунитарные гладкомышечные клетки слишком малы для того чтобы генерировать потенциал действия. (Когда потенциал действия возникает на мембране висцеральной (унитарной) гладкой мышцы, то от 30 до 40 гладкомышечных клеток должны деполяризоваться одновременно до того момента, когда потенциал действия будет способен самостоятельно растпространяться вдоль мембраны гладкой мышцы. В мультиунитарной гладкой мышце не возникает потенциал действия, а локальная деполяризация, вызываемая высвобождением медиатора, способна к электроническому распространению. Особенности актомиозинованого взаимодействия. В гладкой мышце движение актомиозиновых мостиков является более медленным процессом, по сравнению с поперечнополосатой мышцей. Однако время, в течение которого головки миозиновых молекул остаются прикрепленными к актину оказывается более длительным. Причиной столь медленного движения актомиозиновых мостиков гладкомышечных клеток является более низкая АТФ-азная активность головок их миозиновых молекул. Поэтому распад молекул АТФ и высвобождение энергии, необходимой для обеспечения движения актомиозиновых мостиков происходит не так быстро как в поперечнополосатой мышечной ткани. Это можно понять, если представить, что одна молекула АТФ необходима для одного движения актомиозинового мостика независимо от продолжительности данного движения. Экономичность энерготрат в гладкой мышце является чрезвычайно важным в общем потреблении организмом энергии, так как, кровеносные сосуды, тонкий кишечник, мочевой пузырь, желчный пузырь и другие внутренние органы постоянно находятся в тонусе. Особенность электромеханического сопряжения. Продолжительность сокращения гладких мышц может варьировать от 0,2 до 30 сек. Сокращение типичной гладкой мышцы начинается спустя от 50 до 100 мсек после начала ее возбуждения, достигая своего максимума через 0,5 сек, а затем угасает в течение последующих 1-2 сек. Таким образом, продолжительность сокращения составляет 1-3 сек, что является в 30 раз более длительным, чем в поперечнополосатой мышце. Возникновение сокращения в гладкомышечных клетках в ответ на увеличение внутриклеточной концентрации ионов кальция – электромеханическое сопряжение осуществляется гораздо медленнее, чем в поперечнополосатой мышце. Механизм электромеханического сопряжения в гладкой мышце отличается от поперечнополосатой или сердечной мышцы. В гладкой мышце появление на сарколемме потенциала действия активирует фосфолипазу С и появление инозитол-3-фосфата, который связывается со специфическим для него рецептором, расположенным на кальциевом канале терминальной цистерны СПР. Это приводит к открытию этих каналов и выходу кальция из цистерны СПР. Механизм сокращения гладких мышц Несмотря на то, что скелетные мышечные волокна сокращаются быстро, для большинства гладких мышц характерно длительное тоническое сокращение, иногда достигающее часов или даже дней. Можно предположить, что механизм сокращения гладкой мышцы отличается от поперечнополосатой мышцы. Подобно скелетной мышце для начала мышечного сокращения гладкомышечных волокон необходимы ионы кальция. Увеличение внутриклеточной концентрации ионов кальция может происходить при раздражении нервных волокон, под влиянием гормонов, растяжения гладкой мышцы или даже в результате изменения химического окружения мышечного волокна. В гладкой мышце движение поперечных актомиозиновых мостиков, лежащее в основе сокращения, начинается благодаря кальций-зависимому процессу фосфорилирования головок миозиновых молекул. Ионы кальция могут поступать в клетку несколькими путями. • Под влиянием медиаторов. При взаимодействии медиатора с расположенным на поверхности гладкомышечной клетки соответствующим рецептором, происходит открытие рецептор-активируемого Са++ канала и вход ионов кальция внутрь клетки. • Через потенциал-зависимые каналы, открывающиеся при изменении разности потенциалов на мембране гладкомышечной клетки. Ионы кальция могут поступать в клетку через потенциал-зависимые кальциевые каналы, которые открывается в мембране гладкомышечных клеток при появлении на ней потенциала действия. • Источником ионов кальция может быть саркоплазматический ретикулум. В мембране саркоплазматического ретикулума есть каналы, которые активируются (открываются) инозитолтрифосфатом (IP3) и поэтому получили название IP3-рецепторов. Это название позволяет отличать их от рианодиновых рецепторов, обнаруженных в саркоплазматическом ретикулуме поперечнополосатых мышц. Механизм длительно удерживаемого укорочения («latch»-механизм). «Мостики на замке». Дефосфорилированые поперечные мостики, но оставшиеся прикрепленными к актину получили название мостики на замке. Это позволяет гладкой мышце поддерживать тонус при минимальных энергетических затратах и связано с тем, что данные мостики не циклируют и поэтому не требуют большого количества энергии АТФ. Подобное явление в значительно меньшей степени имеет место и в поперечнополосатой скелетной мышце, и также не требует большого числа нервных импульсов и концентрации гормонов. |