Папоротники. курсовая папоротники. 1 Эволюция папоротников
 Скачать 229.2 Kb. Скачать 229.2 Kb.
|

|
Содержание Введение Папоротники – Polypodiophyta – древняя группа, класс высших архегониальных растений, связанная по своему происхождению с псилофитами. Появились впервые в середине девонского периода палеозойской эры. Один из самых многовидовых отделов, причем таксономическое разнообразие проявляется уже в середине каменноугольного периода и сохраняется до наших дней. В настоящее время на Земле существует около 10 000 видов отдела. Для истории планеты именно папоротники имели важное эволюционное значение: все наиболее совершенные и многочисленные высшие растения (голосеменные и покрытосеменные) – их потомки. [stepanov_n_v_i_dr_botanika_konspekt_lektsiy] Папоротники относятся к числу наиболее древних групп высших растений. Их ископаемые остатки богато представлены в отложениях, датируемых каменноугольным периодом, хотя отдельные виды известны с девона. В настоящее время это наиболее многочисленная группа среди споровых сосудистых растений. Они насчитывают более 12 000 видов из 300 родов. Распространены папоротники практически по всему земному шару, начиная от пустынь и арктических тундр до болот и тропических лесов. Две трети видового разнообразия папоротников сосредоточены в тропических широтах, остальные населяют умеренные области Земли. Папоротники представлены многочисленными жизненными формами - от тропических древовидных форм, достигающих высоты 25 м и диаметра ствола 50 см, до крошечных растений длиной всего лишь несколько миллиметров. [https://ido.tsu.ru/other_res/hischool/botanika2/3h.html - Рудая Н.А. Ботаника: систематика растений, Томск – 2004] 1 Эволюция папоротников Самые древние сосудистые растения на Земле – риниофиты. Они появились в силурийском периоде палеозойской эры, около 440 млн. лет назад и росли в прибрежной зоне. Настоящих корней они еще не имели, в почве находился горизонтальный побег, от которого поднимались вверх вертикальные, дихотомически ветвящиеся оси, многие из которых заканчивались спорангиями. Все риниофиты были равноспоровыми растениями. Листья еще отсутствовали, роль корней выполняли ризоиды. Но это уже были сосудистые растения, у них уже сформировалась ксилема, проводящая воду вверх по стеблю и флоэма, проводящая органические вещества, окружала центральный тяж ксилемы. Центральный проводящий пучок был окружен механической тканью и клетками коры, снаружи уже была покровная ткань – эпидерма, имеющая устьица. Механическая, проводящая и покровная ткани позволили растениям приспособиться к жизни в воздушной среде и начать освоение суши. Дальнейшее освоение суши сопровождалось появлением корней и листьев. От одной из групп риниофитов (зостерофиллофитов) произошли плауновидные, причем листья у них образовывались как уплощенные боковые стебли с единственной жилкой (проводящим пучком), такие листья называют микрофиллами. Папоротниковидные и, возможно, хвощевидные растения произошли от другой группы риниофитов – псилофитов. Листья у них образовывались из системы боковых разветвленных уплощенных побегов, называются мегафиллами и имеют сложную систему жилок. Расцвет папоротников произошел около 358 млн лет назад и длился примерно 65 млн лет, в честь папоротников назван период палеозойской эры - карбон, или каменноугольный период, который длился указанный промежуток времени. Именно папоротникам принадлежит активная роль в углеобразовании: в карбоне леса заселяли древовидные папоротники, достигавшие 40 и более метров в высоту. В науке считается, что папоротникообразные появились на земле в период среднего девона. Их численность и разнообразие просто поражают. Папоротники начинают разрастаться, появляются древовидные формы. Более сложная структура позволяет этим видам адаптироваться к жизни на суше и придает им необходимую для дальнейшего роста жесткость. В результате значительного накопления остатков этих растений стали появляться залежи торфа, который со временем преобразовался в каменный уголь. В последующие столетия появились благоприятные условия и влажная среда для разрастания папоротниковидных. Археологи часто находят в залежах угля на поверхности этой породы рисунки листьев папоротников и отмечают появление новых форм флоры. В каменноугольный период, 360 млн. лет назад, доминировали хвощи. Из их окаменевших останков образовалась большая часть угля. Затем они постепенно были вытеснены другими видами. Папоротники нашего времени растут в богатых влагой тропических лесах. Встречаются травянистые и древовидные растения, а также лианы, которые схожи по размерам с древними папоротниками. [https://obrazovanie.guru/nauka/biologiya/paporotnikoobraznye-rasteniya.html obrazovanie.guru] Важнейшее преимущество папоротникообразных еще и в том, что в жизненном цикле полностью доминирует диплоидный спорофит. Происходит накопление мутаций и их комбинации в потомстве попадают под контроль естественного отбора. Проводящая система папоротникообразных представлена пока еще не сосудами (трахеями), а трахеидами, и во флоэме ситовидные клетки без клеток-спутниц, ситовидные трубки появятся позже, у цветковых. Гаметофиты имеют небольшие размеры, развиваются независимо от спорофита и образует яйцеклетки и сперматозоиды, для слияния которых нужна вода. Таким образом, папоротникообразные – «земноводные растения», спорофиты приспособлены к жизни на суше, а для развития гаметофитов еще нужна вода. [https://sbio.info/lections/rasteniya/paporotnikoobraznye] Наивысшего расцвета папоротникообразные достигали в каменноугольный период, когда папоротникообразные занимали на суше огромные территории и были широко распространены по всему земному шару. Папоротникообразные были представлены большим разнообразием форм, в том числе мощными древовидными папоротниками. В настоящее время папоротникообразные сохранились главным образом в виде травянистых растений. Древовидные папоротники произрастают в небольшом количестве только в тропиках. Очень не многие представители этого отдела являются водными растениями. В цикле развития папоротникообразных явно стал преобладать спорофит, который хорошо развит; гаметофит же морфологически, наоборот, сильно редуцирован, особенно у разноспоровых папоротников. Дальнейшая эволюция папоротникообразных шла именно в направлении ещё большей редукции гаметофита, спорофит же развивался очень сильно, что способствовало появлению семенных папоротников, у которых оплодотворение яйцеклетки и дальнейшее развитие зиготы и зародыша происходило непосредственно на спорофите. Семенные папоротники в настощее время вымерли, они являются родоначальниками более высоко организованных растений - голосеменных. В настоящее время папоротникообразные не имеют существенного практического значения, их используют как лекарственные (глистогонные), декоративные, редко как пищевые (в пищу употребляют молодые побеги). В далеком же прошлом папоротники совместно с другими растениями образовывали огромные леса, именно благодаря им образовывались залежи каменного угля. [В.В.Суворов, «Ботаника с основами геоботаники»- Л.:1971г.] 2 Классификация папоротников За последние годы систематика папоротников достигла больших успехов. Это объясняется прежде всего значительным прогрессом исследований в области сравнительной морфологии современных, так называемых ныне живущих, и вымерших папоротников. Сделано очень много, в частности, в изучении устьиц, спор и гаметофитов. Накоплен богатый материал по цитологии папоротников. Все это позволило пересмотреть старые схемы классификации папоротников и подойти значительно ближе к выяснению родственных взаимоотношений между ныне живущими группами папоротников и направлением их эволюции. Синтез всех этих данных позволяет строить классификацию и филогению папоротников на гораздо более надежном фундаменте, чем это было, например, в 1935 г., когда известный английский птеридолог Ф. Бауэр предложил новый вариант своей филогенетической классификации. И хотя среди птеридологов все еще имеются большие разногласия, в частности об объеме высших таксономических единиц папоротников, общая картина эволюции значительно прояснилась. На современном уровне отдел папоротниковидных можно подразделить на следующие 7 классов. [ Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974. ] 2.1 Современные папоротники Известно большое количество ископаемых и современных форм из числа папоротниковидных. Современных представителей отдела относят к трем классам: Ophioglossida, Marattiopsida и Polypodiopsida. Из них только виды классов Ophioglossida и Polypodiopsida представленны во флоре России. Всего на земном шаре обитает около 12 000 видов папоротников. [uranov_a_a_sistematika_rasteniy] 2.1.1 Класс Marattiopsida Папоротники, появившиеся в середине каменноугольного периода палеозойской эры, и одна из немногих древних групп, сохранившихся до настоящего времени (рис 1). 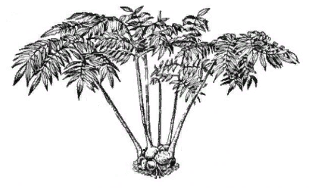 Рисунок 1 - Ангиоптерис (Angiopteris evecta) — общий вид растения Мараттиопсиды берут начало от зигоптерисовых папоротников, с которыми их сближают многие архаические черты: эуспорангиатность, наличие афлебиев, отсутствие приспособлений для раскрытия спорангиев или примитивное кольцо. Для современных мараттиевых характерны срастание спорангиев в синангии и их ламинальное расположение. Жизненную форму можно, вероятно, считать полудревовидной (редукция древовидности). Синтелом в виде дорсо-вентрального корневища или клубневидного ствола отличается мясистостью. В молодых синтеломах имеется протостела, которая в дальнейшем преобразуется в амфифлойную сифоностелу. В вегетативных органах мараттиевых развиты большие лизигенные ходы. Листья большинства мараттиопсид перистые, но бывают пальчатые и цельные. Молодые листья всегда спирально закрученные. Спорангии довольно крупные, расположенные на нижней стороне листьев и собраны с сорусы или синангии. Число спор в каждом спорангии очень велико. Равноспоровые растения. Спорангии довольно крупные, толстостенные и снабжены устьицами. Расположенные на нижней (абаксиальной) стороне зеленых листьев вдоль жилок спорангии образуют сорусы или срослись в различного рода синапгии. Покрывальце отсутствует. Зрелые спорангии раскрываются посредством щелей или пор, причем у некоторых родов (ангиоптерис, макроглоссум — Macroglossum, архангиоптерис — Archangiopteris) имеются примитивные кольца, напоминающие по своему строению кольца некоторых зигонтеридопсид. Споры трилетные или монолетные. Число их в каждом спорангии очень велико (от 1450 до 7500). Гаметофиты надземные, темно-зеленые, мясистые, дорсивентральные, сердцевидные или продолговатые, долговечные. Антеридии большие и погруженные, как у офиоглоссопсид, и развиваются сходным образом. Архегонии сходны с архегониями офиоглоссопсид, но шейки их более короткие и широкие. В класс входит один порядок — мараттиевые (Marattiales), который состоит из одного современного семейства Marattiaceae, иногда разделяемого на три семейства — Marattiaceae, Christenseniaceae и Danaeaceae. В двух последних семействах но одному роду. Представители - ангиоптерис (Angiopteris), мараттия (Marattia). [Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.] [https://ido.tsu.ru/other_res/hischool/botanika2/3h.html - Рудая Н.А. Ботаника: систематика растений, Томск – 2004] [uranov_a_a_sistematika_rasteniy] 2.1.2 Класс Polypodiopsida Появился, очевидно, в первой половине пермского периода палеозойской эры около 270 млн лет назад. Считается современным. Предковыми для полиподиевых папоротников являются ботриоптеридопсиды. Представители класса наиболее сложно организованы по сравнению с современными папоротниками и характеризуются лептоспорангиальностью, различными типами вай, самыми совершенными видами спорангиев, различными типами спороношений, высокоорганизованной проводящей системой. [uranov_a_a_sistematika_rasteniy] Современные папоротники. Геологическая история восходит к карбону. Многолетние и редко однолетние растения, варьируют от мелких форм до достаточно крупных. Стебли представлены стволами (у древесных форм весьма крупных размеров) или превращены в корневища. Листья различных типов по размерам, форме, жилкованию, плотности. Спорангии, как правило, собраны в сорусы. Равноспоровые и разноспоровые растения. Гаметофиты у большинства полиподиопсид зеленые, наземные, обоеполые, но у представителей разноспоровых - раздельнополые и сильно редуцированные. [https://ido.tsu.ru/other_res/hischool/botanika2/3h.html - Рудая Н.А. Ботаника: систематика растений, Томск – 2004] Включает несколько подклассов. Osmundidae – группа примитивных папоротников, занимающих промежуточное положение между эуспорангиатными и лептоспорангиатными таксонами. Достаточно обособлена от всех подклассов полиподиопсид, очевидно вследствие ранней дивергенции от остальных таксонов полиподиевых папоротников. Спорангии крупные, без кольца. В синтеломных корневищах относительно сильно развита кора при тонкой стеле. Тип проводящей системы – диктиостела; при этом флоэма окружает «сетчатую» флоэму в виде сплошного цилиндра. Представители известны по ископаемым останкам пермского периода. Schizeidae – своеобразная, достаточно древняя группа папоротников. Известна с каменноугольного периода палеозойской эры. Представители подкласса характеризуются комплексом достаточно примитивных черт: одиночные спорангии расположены по краю пластинки, созревают одновременно (симультанный тип); верхушечное поперечное кольцо, способствующее вскрытию спорангия; трехлучевые споры, дихотомическое жилкование в сегментах вай; диморфизм в строении генеративных и вегетативных вай или их частей. Порядок птерисовых считается достаточно специализированным. Спорангии, вперемешку с парафизами, располагаются непрерывной или прерывистой линией вдоль края сегмента пластинки вайи, образуя так называемый ценосорус. Спорангии при этом прикрыты подвернутым краем пластинки. Индузий отсутствует. Созревание спорангиев неопределенное. Кольцо продольное. Споры трехлучевые или однолучевые (редко). Жилкование дихотомическое. Проводящая система – соленостела. Корневищеподобные синтеломы покрыты чешуями, реже волосками. Hymenophyllidae – достаточно разнородная в отношении степени примитивности либо эволюционной продвинутости группа папоротников. Наблюдается различный характер развития синтелома: от развитого плагиотропного до редуцированного ортотропного у некоторых «короткокорневищных» папоротников. Спорангии собраны в градатные сорусы и имеют поперечное, косое или продольное кольцо. Созревание спорангиев – базипетальное. Индузий обычно хорошо развит. Споры трех– или однолучевые. Происходят, вероятно, от вымерших ботриоптерисовых. Известные представители следующие. Pteridium aquilinum (Орляк обыкновенный). Один из самых распространенных папоротников Земли (рис 2). Встречается на всех континентах, кроме Антарктиды. Чрезвычайно изменчив и в разных регионах представлен особыми географическими расами – подвидами. Произрастает в светлых смешанных лесах, а также в сосняках, темнохвойной горной тайге, на лесных полянах, по берегам рек, на остепненных лугах и др. 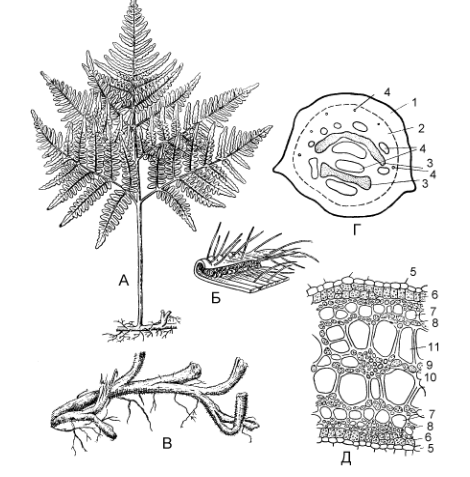 Рисунок 2 - Папоротник-орляк (Pteridium aquiliunum) А – общий вид; Б – часть листочка с сорусами; В – часть корневища; Г – поперечный разрез корневища (схема); Д – часть пучка на поперечном разрезе (большое увеличение): 1 – наружная кора, 2 – внутренняя кора, 3 – полукольцо механической ткани, 4 – проводящие пучки, 5 – эндодерма, 6 – перицикл, 7 – ситовидные трубки, 8 – лубяная паренхима, 9 – древесинная паренхима, 10 – первичная ксилема, 11 - ксилема Для орляка характерны длинные подземные синтеломы («корневища»), черно-бурые. Диаметр – 1 – 1,5 см. Глубина их залегания – от 5 до 60 см. Возраст синтеломов, определенный по оставшимся, неразложившимся частям, составляет более 50–70 лет. Вайи высотой 30–200 см, а в тропиках – до 4 метров. Примерно половину высоты вайи занимает черешок, а еще половину – крупная треугольная или яйцевидная пластинка. Молодые вайи более или менее густо покрыты коричневыми или белыми волосками; имеют промышленное пищевое значение. В Японии выращивается в культуре. В некоторых регионах – злостный сорняк, который засоряет пастбища, ядовит для скота. Используется в народной медицине, имеет техническое применение. Dryopteris filix-mas (Щитовник мужской). Вайи продолговатоланцетные, короткочерешковые, дваждыперистые. Произрастает в широколиственных, смешанных и темнохвойных лесах. Встречается в Европе, на Кавказе, в Южной Сибири, Средней Азии, Северной Америке. Сибирские местонахождения мужского щитовника представлены остаточными фрагментами ареала, изолированными от основных мест произрастания вида обширными разрывами (дизъюнкциями). В Сибири Dryopteris filix-mas является неморальным реликтом третичного периода кайнозойской эры. Polypodiidae – один из «молодых» в филогенетическом отношении подклассов. Лептоспорангиатные. Представители в значительной степени специализированы. Спорангии – в сорусах. Характерно отсутствие (за редкими исключениями) индузия. По своему происхождению связаны, вероятно, с вымершими представителями предыдущего подкласса. Характерные представители следующие. Polypodium (Многоножка). Насчитывает 75 видов, являющихся некрупными эпифитами или наземными формами с перисторассеченными пластинками вай. Относится к типовому подсемейству Polypodioideae. Распространены в тропической, субтропической и умеренной зонах. В России встречаются 5 видов, из которых два достаточно широко представлены в Южной Сибири. Polypodium sibiricum (многоножка сибирская) распространен к востоку от Енисея. Имеет синтелом, покрытый темно-бурыми, почти черными чешуйками, более мелкие размеры вай (до 25 см в длину, 2–4 см в ширину), сорусы, несколько сдвинутые к краю пластинки. Другой вид – Polypodium vulgare (многоножка обыкновенная) – встречается к западу от Енисея (до Западной Европы и Северной Америки). Синтелом покрыт светлобурыми чешуйками, вайи относительно более крупные (до 35 см в длину, 4–6 см в ширину), сорусы расположены в середине сегмента, «корневище» имеет сладкий вкус. Сибирские виды распространены в горных районах, где встречаются в лесной полосе, изредка – в высокогорьях. Произрастают на тенистых скалах, крупных мшистых валунах, иногда на колоднике, очень редко в основании стволов деревьев. Marsileidae – водные и земноводные разноспоровые папоротники. Известны с конца мелового периода мезозойской эры. Этот подкласс в систематическом отношении имеет изолированное положение и, по-видимому, происходит от циатейных папоротников, представителям которого близок по данным геномного анализа. Иногда сближается со схизейными папоротниками. Сорусы – в спорокарпиях, представляющих собой видоизмененный предпобег. Спорокарпии твердые сидячие или на ножках. В каждом сорусе находятся мега– и микроспорангии. Многолетники с тонким, ползучим плагиотропным синтеломом и двурядно расположенными на нем вайями. Вайи специфической формы: нитевидные или похожие на лист клевера, но «четырехлисточковые». В подклассе – единственный порядок и семейство, три рода и около 80 видов, распространенных преимущественно в Восточном полушарии. Сальвиниевые – Salviniidae – водные разноспоровые папоротники, конвергентные по отношению к марсилиевым. Известны с начала мелового периода. Произошли, очевидно, от гименофилловых папоротников. Растения свободноплавающие, лишь изредка могут развиваться на сырой почве. Микро– и мегаспорангии находятся в раздельных сорусах. Каждый сорус полностью покрыт индузием, образуя спорокарпий. В подклассе единственный порядок и два семейства. Это водные однолетние растения с тонкими синтеломами. Вайи – по трое в мутовках, видоизмененные и диморфные: две из них плавают на поверхности воды и имеют цельные, неперистые пластинки; одна погружена в воду и похожа на корни. Сорусы на ножках, полностью скрыты индузием. Микроспорангии содержат 9–14 микроспор; мегаспорангии с одной–двумя мегаспорами. В семействе один род. [stepanov_n_v_i_dr_botanika_konspekt_lektsiy][ uranov_a_a_sistematika_rasteniy] 2.1.3 Ophioglossopsida Ужовниковые – своеобразная группа современных растений, мало похожих на папоротники (рис). Современный отдел. Произошли, очевидно, от равноспоровых аневрофитопсид в палеозое. Есть три современных рода и более 80 видов. Многолетние травянистые растения, как правило, небольших размеров. Небольшие или средних размеров многолетние растения, обычно наземные, редко эпифиты (некоторые тропические виды ужовника — Ophioglossum). 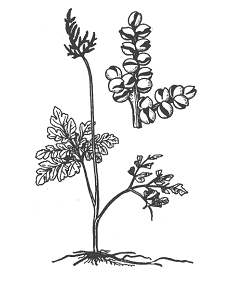 Рисунок - Гроздовник (Botrychium lunaria) – общий вид Синтелом, корневищеподобный, укороченный, имеет проводящую систему сифоностелического или диктиостелического типов. Корневище короткое, простое. У крупных офиоглоссид имеется активно функционирующий камбий и развивается вторичная ксилема, что отличает их от других современных представителей папоротников. Чешуи на корневищах отсутствуют. Проводящая система корневища представляет собой эктофлойную сифоностелу или диктиостелу, но у некоторых представителей (гроздовник — Botrychium, гельминтостахис - Helminthostachys, и ряд видов ужовника) молодое корневище имеет протостелическое строение. Ксилема гельминтостахиса экзархная, но у остальных родов она эндархная, что несколько необычно для папоротников. У более крупных гроздовников имеется активно функционирующий камбий, причем у гроздовника вирджинского (Botrychium virginianum) вторичная ксилема хорошо развита и имеет лучи. Эта особенность отличает офиоглоссопсиды почти от всех остальных современных папоротников и сближает их с палеозойскими аневрофитопсидами. Более того, у гельминтостахиса и гроздовника окаймленные поры трахеид округлые или овальные и с явственным торусом, как у многих голосеменных. Листья отличаются от листьев других современных папоротников отсутствием улиткообразного закручивания в молодости, наличием особых влагалищных прилистникоподобпых образований при основании, а также обычно мягкостью. Подобно корневищам они обнаруживают иногда признаки дихотомического ветвления. Характерна диморфность листьев. Некоторые виды способны к вторичному росту (остаточному) и во вторичной ксилеме; имеют лучи. Трахеиды – окаймленнопоровые с торусом, как у голосеменных. Это равноспоровые эуспорангиатные растения. Спорангии довольно крупные, содержат большое количество спор (1500–15000). Вайи в эмбриональном состоянии не свернуты улиткой. Гаметофиты мясистые, цилиндрические, дихотомически или неправильно ветвящиеся, многолетние, обычно подземные, микоризные; эндомикосимбионты. гаметофит живет 10-20 лет, обладая хорошо развитой верхушечной меристемой и также способен дихотомически ветвиться. [stepanov_n_v_i_dr_botanika_konspekt_lektsiy] [https://coollib.net/b.usr/Aleksandr_Lvovich_Ivanov_Evolyutsiya_i_filogeniya_rasteniy.pdf] Спорофиты мясистые, лишены механической ткани. Растения равноспоровые. Спорангии довольно крупные, содержат большое число спор (от 1500 до 15 000), имеют массивные многослойные стенки и снабжены устьицами. Они лишены кольца и раскрываются морфологически верхушечной щелью. Споры трилетные, вначале тетраэдрические, но затем становятся почти шаровидными. У некоторых видов в спорах наблюдаются бледные хлоропласты. В классе один порядок ужовниковые (Ophioglossales), одно семейство ужовниковые (Ophioglossaceae) с тремя подсемействами — Botrychioideae, Helminthostachydoideae и Ophioglossoideae. Основные представители ужовник (Ophioglossum) и гроздовник (Botrychium). Все Ужовниковые – облигатные (обязательные) микотрофы, но степень зависимости от микоризы различна у разных видов. Гельминтостахис и эпифитные ужовники являются вечнозелеными растениями, но максимум образования новых листьев и период спороношения у них приходятся на определенное время года. У наземных ужовников умеренной зоны листья летнезеленые; в пределах рода Гроздовник есть виды летнезеленые и «зимнезеленые». Хотя хозяйственного значения Ужовниковые не имеют, эта своеобразная и малочисленная группа заслуживает бережного отношения к себе, а многие виды уже давно нуждаются в охране. Так, вид Botrychium simplex занесен в Красную книгу РФ. [https://ido.tsu.ru/other_res/hischool/botanika2/3h.html - Рудая Н.А. Ботаника: систематика растений, Томск – 2004] [ Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.] 2.2 Древнейшие папоротники 2.2.1 Cladoxylopsida Наиболее примитивный и древний класс папоротников. Появились в середине девонского периода палеозойской эры, т. е. около 370 млн лет назад. Исчезли в начале каменноугольного периода, просуществовав всего 40–50 млн лет. Несмотря на кратковременное существование, дали начало по меньшей мере двум крупным филогенетическим ветвям, что имело значительное эволюционное значение. Финалом одного из направлений сегодня являются современные папоротники, другого – два существующих ныне отдела высших растений, т. е. голосеменные и покрытосеменные. Произошли кладоксилопсиды от псилофитов. Кладоксилеевые папоротники и внешне очень похожи на них: отсутствуют уплощения осевых органов; как следствие, у них – синтеломный уровень организации; имеются крупные терминальные спорангии; характерны более или менее сильные разветвления; иногда, кроме ортотропных, присутствуют и плагиотропные синтеломы. Особенностями этой группы папоротников являются появление более или менее кладифицированных микротеломов, а главное – сложно расчлененная проводящая система осевых частей. Ксилема экзархная. В районе протоксилемы присутствуют периферические петли или протоксилемные лакуны. Хотя представители девонского рода псевдоспорохнус (Pseudosporochrius) достигали, вероятно, в высоту 2 м, среди кладоксилопсид не было настоящих древесных форм, если не считать один вид кладоксилона (Cladoxylon nodosus), который имел, по-видимому, облик маленького деревца. Листья были дихотомически разветвленные. Проводящая система представляла собой довольно своеобразную рассеченную актиностелу, многочисленные меристелы которой составляли вдоль по длине стебля анастомозирующую систему. Ксилема состояла из лестничных и точечных трахеид. Спорангии были верхушечные и лишены каких-либо специальных приспособлений для раскрывания. Кладоксилопсиды были равноспоровыми растениями. Они представляли собой, вероятно, слепую ветвь эволюции. Важнейшие представители следующие. Pseudosporochnus nodosus (Псевдоспорохнус узловатый). Ранее считался псилофитом. Встречался в середине девона в Западной Европе. Небольшое, высотой около одного метра, растение было представлено только ортотропными осями. Неразветвленный ствол в верхней части увенчан пучковидной «мутовкой» ветвей, в нижней части – пучок корней. Ветви сильно разветвленные. Конечные веточки были частично стерильными, частично – с двумя парными спорангиями. Проводящая система напоминает сильно расчлененную актиностелу либо «упорядоченную» плектостелу. В центре протоксилемы проходит тяж тонкостенных паренхимных клеток. Благодаря этому образуются периферические петли. Cladoxylon mirabile (Кладоксилон удивительный). Встречался в начале карбона в Западной Европе. Достоверно к этому роду можно отнести только окаменелые фрагменты ортотропных осей. Близок к псевдоспорохнусу. Проводящая система – полистелического типа. Расположение лент ксилемы неупорядоченное. Появляется вторичная древесина. Диаметр стволов – до 4 см. На поперечном срезе присутствуют листовые следы, направляющиеся в афлебии. Название составлено из латинских слов, обозначающих «ветка» + «древесина», и указывает на разветвление ксилемных лент на поперечном разрезе. Protohyenia janovii (Протогиения Янова). Встречалась в начале девона. Известна по ископаемым останкам с правого берега Енисея у г.Красноярска (Торгашино). Внешним обликом напоминает предыдущий таксон, но генеративные ветви, как у псилофитов. Спорангии раскрывались продольно с помощью верхушечной щели. 2.2.2 Zygopteridopsida Вымерший в середине пермского периода палеозойской эры таксон. Первые растения появились в конце девона. Предковой группой, очевидно, являлись кладоксилеевые папоротники. Класс объединяет довольно разнообразных представителей, часто контрастных по степени эволюционной специализированности и приспособленности. Это типичные эуспорангиатные растения. Спорангии располагались одиночно или группами, терминально (в особых по морфологической природе органах – в спорокладах, отделенных от вегетативных частей) или на нижней поверхности пластинки вайи. На стенках спорангия появляется дифференцированная группа клеток, способствующая их вскрытию. Споры трехлучевые. Характерны различная форма ксилемы в осевых органах (C-, V-, W-, I-, H- или Xобразная), а также протостела или актиностела. У некоторых представителей в ксилеме развиваются периферические петли, как у кладоксилеевых папоротников. Зигоптерисовые папоротники – первое эволюционное звено в направлении всех современных папоротников кроме ужовниковых. Именно в рамках данного таксона у «папоротниковой» линии эволюции впервые, в начале карбона, появляются листовидные образования – вайи (предпобеги). Необычен тот факт, что в другой эволюционной линии (проголосеменные и голосеменные) вайи появились значительно раньше. Для зигоптерисовых характерно наличие особых дорсо-вентральных органов – филлофоров, промежуточных по своей природе между синтеломными корневищами и рахисами плосковеток и предпобегов. Филлофоры ответвлялись от плагиотропных синтеломов и росли разным образом. Так же, как и у синтелома, их рост был неограниченный. На них развивались перья, располагавшиеся либо в два, либо в четыре ряда. По мпению многих исследователей, от зигоптеридопсид берут начало многие типичные папоротники (в частности, осмундовые, схизейные и глейхениевые). В этот класс входит порядок зигоптерисовые (Zygopteridales), содержащий несколько семейств (в том числе зигоптерисовые — Zygopteridaceae, ботриоптерисовые — Botryopteridaceae и ставроптерисовые — Stauropteridaceae). Вполне возможно, что зигоптеридопеиды представляют собой не вполне естественную группу. Класс включает единственный порядок. Основным представителем является Зигоптерис (Zygopteris). [?] [Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.] 2.2.3 Aneurophytopsida Это очень древняя группа, геологическая история которой известна с конца раннего до середины позднего девона (400—375 млн. лет назед). Наиболее примитивные представители этого класса (особенно среднедевонский род птилофитон — Ptilophyton, известный также под названием Protopteridium) имеют еще много общего с риниофитами и представляют собой, в сущности, промежуточные формы между ними и папоротниками. Предполагается, что они произошли от представителей порядка псилофитовых (Psilophytales). Растения, относящиеся к этому классу, еще не имели настоящих листьев. Лишь у птилофитона конечные веточки были уже уплощенные, листоподобные. Это были своего рода «плосковетки» — первая стадия в возникновении настоящих, плоских, дорсивентральных листьев. Но подобно листьям настоящих папоротников конечные веточки аневрофитопсид были в молодости улиткообразно закручены (рис. 93). Внутреннее строение ветвей всех порядков еще очень примитивное, всегда протостелическое. Первичная ксилема мезархная. Почти у всех представителей класса происходило образование вторичной ксилемы, а у некоторых из них — довольпо сложно устроенной вторичной флоэмы. Трахеиды вторичной ксилемы были точечные, т. е. с округлыми окаймленными порами. Спорангии были относительно крупные, верхушечные, одиночные или собранные в пучки. Споры одинаковые, с трехлучевым тетрадным рубцом. Известный американский ботаник Д. Бирхорст (1971) включает в свой класс аневрофитопсид (название класса было предложено им) также представителей следующего далее класса археоптеридопсид. Однако в настоящее время представляется более правильной точка зрения русского палеоботаника Н. С. Снигиревской, которая предлагает разделить аневрофитопсиды Бирхорста на два самостоятельных класса — аневрофитопсиды в более узком смысле и археоптеридопсиды. В класс аневрофитопсид входит порядок аневрофитовых (Aneurophytales), а также ряд растений, родство которых еще не вполне точно установлено. Наиболее примитивный среднедевонский род птилофитон (Ptilophyton). https://ido.tsu.ru/other_res/hischool/botanika2/3h.html - Рудая Н.А. Ботаника: систематика растений, Томск – 2004 Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974. [uranov_a_a_sistematika_rasteniy] 2.2.4 Archaeopteridopsida Представители этого класса известны из отложений конца среднего девона — начала карбона (390—360 млн. лет назад). От аневрофитопсид, с которыми их обычно объединяют, они хорошо отличаются наличием настоящих листьев с цельной, вильчато, пальчато или перисторассеченной пластинкой. Но вот что очень интересно и в эволюционном отношении очень важно: эти, казалось бы, столь сходные с листьями папоротников настоящие листья, в молодости не бывали улиткообразно закрученными. Этим они наноминают листья современных офиоглоссопсид (ужовниковых). Строение ветвей сифоностелическое, а не протостелическое, как у аневрофитопсид. Первичная ксилема состояла из кольца мезархных пучков, сливавшихся при образовании вторичной ксилемы в цилиндрический тяж. Взрослые растения имели древовидный облик, отдаленно напоминая современные хвойные (рис. 94). Вторичная ксилема достигала большой мощности, т. е. древесина была пикноксильного типа. Трахеиды вторичной ксилемы были точечные, чем археоптеридопсиды отличаются от всех остальных папоротниковидных, за исключением аневрофитопсид и офиоглоссопсид. Спорангии крупные, расположенные одно- или двурядно на отдельных перисторассеченных спорофиллах, чередующихся со стерильными листьями. Таким образом, по расположению спорангиев археоптеридопсиды резко отличаются от аневрофитопсид, что еще более подтверждает необходимость рассматривать их как два самостоятельных класса. Кроме того, у многих представителей археоптеридонсид известна гетероснория (разноспоровость). В классе один порядок — археоптерисовые (Агchaeopteridales). Основываясь на наличии ряда общих черт с голосеменными (особенно на наличии вторичной древесины, которая у археоптериса вполне голосеменного типа, а также на разноспоровости ряда представителей), американский палеоботаник Ч. Бек (1960) назвал этот класс (понимаемый им шире, чем здесь) «прогимноспермами» (Progymnospermopsida). Однако Д. Бирхорст (1971) совершенно правильно указывал на то, что название таксона не должно отражать филогенетическую интерпретацию, и поэтому предложил более правильное с точки зрения ботанической номенклатуры название — аневрофитопсиды. Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974. Заключение Более 400 000 видов флоры известны сегодня, и все они произошли от нескольких древних морских растений. Виды, исчезнувшие с лица Земли, не входят в это число, поскольку к меняющимся условиям на Земле адаптироваться они не смогли, или не выдержали конкуренцию со стороны вновь появившихся растений, которые лучше были приспособлены к новой среде обитания. Наивысшего расцвета папоротниковидные достигли в каменноугольный период, когда занимали на суше огромные территории и были широко распространены по всему земному шару. Они были представлены большим разнообразием форм, в том числе мощными древовидными папоротниками. В настоящее время папоротниковидные сохранились главным образом в виде травянистых растений. Древовидные папоротники произрастают в небольшом количестве только в тропиках. Очень немногие представители того отдела являются водными растениями. Папоротниковидные произошли от псилофитовидных, в эволюции их проявляется макрофиллия (крупнолистность). Первичные папоротники не имели развитых листьев. В цикле развития папоротниковидных явно стал преобладать спорофит, который хорошо развит; гаметофит же морфологически, наоборот, сильно редуцирован, особенно у разноспоровых папоротников. Дальнейшая эволюция папоротниковидных шла именно в направлении еще большей редукции гаметофита, спорофит же развивался очень сильно, что способствовало появлению семенных папоротников, у которых оплодотворение яйцеклетки и дальнейшее развитие зиготы и зародыша происходили непосредственно на спорофите. Семенные папоротники в настоящее время вымерли, они являются родоначальниками более высоко организованных растений — голосеменных. Отдел папоротниковидные объединяет 7 классов, в которые входят вымершие и ныне живущие папоротники. Аневрофитовые (Aneurophytopsida); Археоптерисовые (Archaeopteridopsida); Кладоксиловые (Cladoxiyopsida); Зигоптерисовые (Zygopteridopsida); Офиоглоссовые (Ophioglossopsida); Мараттиевые (Marattiopsida); Полиподиевые (Polypodiopsida). Представители первых четырех полностью вымерли уже в конце палеозоя. Предполагают, что наиболее примитивные из них – Аневрофитовые – произошли от риниевых, листья их напоминали ассимиляционные ветви. В классах археоптерисовых и зигоптерисовых было много крупных древовидных форм, обладавших вторичным ростом и довольно высоко организованной проводящей системой. Расцвет их пришелся на конец девона и карбон. Именно эти растения в наибольшей степени представлены в каменноугольных отложениях По размерам папоротники варьируют от крупных тропических древовидных форм (выс. до 25 м, диам. ствола до 50 см) до крошечных растений длиной в несколько мм. Стебли папоротников различаются по форме и анатомическому строению. Прямостоячие стволы древовидных папоротников у основания обычно снабжены многочисленными воздушными корнями. придающими им устойчивость. Стебли травянистых папоротников, особенно молодые, покрыты волосками или чешуйками. Листья папоротников, которые часто называют вайями, весьма разнообразны. Их длина колеблется от нескольких миллиметров до 30 м. Обычно лист имеет черешок и пластинку разной формы (простую, перистую, дважды или многократно перистую и т. д.). Листья, как правило, совмещают две функции – фотосинтеза и спороношения, лишь у немногих видов они дифференцированы на фотосинтезирующие (стерильные) и несущие спорангии (фертильные). На определённой стадии развития спорофита начинается спороношение (в биологическом смысле оно аналогично цветению). Спорангии расположены, как правило, на нижней стороне листа или по его краю, у большинства папоротников они сгруппированы в компактные кучки – сорусы. Часто сорусы прикрыты защитными органами – покрывальцами (индузиями). Спорангии одного соруса у разных видов созревают или почти одновременно, или последовательно. При созревании спорангии вскрываются и споры высыпаются. Из множества спор, производимых взрослым папоротником, лишь небольшая их часть попадает в благоприятные условия, прорастает, давая начало половому поколению – гаметофиту, или заростку. Для прорастания спор большинства папоротников, кроме влажности и тепла, нужен свет. У разноспоровых папоротников гаметофит однополый, у равноспоровых – обоеполый. Папоротники являются важным компонентом многих растительных сообществ, особенно в тропических, субтропических, а также северных (преимущественно широколиственных) лесах. Многие папоротники являются индикаторами различных типов почв. Некоторые виды папоротников применяются в медицине как глистогонное средство, для лечения открытых ран, кашля и болезней горла. Виды азолы используются в качестве зеленого удобрения, обогащающего почву азотом. Некоторые папоротники используются в декоративном цветоводстве. Список используемых источников |