|
Ответы к коллоквиуму по физике. 1 Какие процессы называются колебательными Гармонические колебания. Уравнение и график гармонических колебании. Частота колебаний. Укажите на графике амплитуду, период колебания, начальную фазу
№ 25 Липидный бислой, его строение, его толщина. Жидкостно-мозаичная модель мембраны. Основные функции биологических мембран. Диффузия липидных молекул в мембранах. Люминесцентный метод излучения подвижности молекул в мембране (флуоресцентные метки и зонды). Частота перескоков молекул.
Биологические мембраны являются важной частью клетки. Они ограничивают клетку от окружающей среды, защищают ее от вредных внешних воздействий, управляют обменом веществ между клеткой и ее окружением, способствуют генерации электрич. Потенциалов, участвуют в синтезе АТФ в митохондриях. А так же исполняют рецепторную функцию. Так же генерация и распространение биопотенциалов.
Все мембраны построены в основном из белков и липидов. Липидная молекула состоит из 2х частей: несущие электрический заряд головки и длинные хвосты.
Любая мембрана своей структурной основой имеет липидный бислой. Состоящий из двух мономолекулярных пленок липидов, обращенных друг к другу гидрофобными хвостами и контактирующих с окружающей средой полярными головками.
Во всех мембранах бислой выполняет 2 функции:
1)Матричную
2)Барьерную
Является структурной основой для размеения основных рецепторных и ферментных систем клетки, с др. стороны двойной слой липидов является преградой для ионов и водорастворимых молекул.
Жидкостно – Мозаичная модель – (в основе лежит липидная бислойная мембрана) Это липидная основа представляет собой как бы двумерный растворитель, в котором плавают более или менее погруженные белки.
Липиды могут перемещаться в плоскости мембраны (латеральная диффузия) – хаотическое тепловое перемещение молекул липидов и белков в плоскости мембраны. При латер. Диффузии рядом расположенные молекулы липидов скачком меняются местами, и вследствие таких последовательных перескоков из одного места в другое молекула перемещается вдоль поверхности мембраны.
Перемещение молекул по поверхности мембраны клетки за время t определено экспериментально методом флюоресцентных меток – флюоресцирующих молекулярных групп.
Флюоресцентные метки делают флюоресцирующими молекулы, движение которых можно изучать исследуя под микроскопом скорость расплывания по поверхности клетки флюоресцирующего пятна.
Частота перескоков(число перескоков в секунду) молекулы с одного места на другое вследствии латеральной диффузии может быть найдена по формуле
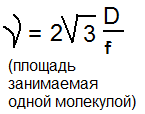
Липиды могут переходить из одного монослоя в другой – ФЛИП-ФЛОП. – дифф. Молекул мембранных фосфолипидов поперек – мембраны.
Скорость перескоков молекул с одной поверхности мембраны на другую определена методом спиновых меток опытах на модельных липидных мембранах – липосомах.
Липидная фаза биологических мембран при физиологических условиях (т-ре, давление) находится в жидком агрегатном состоянии. Это доказано методами фиюоресцентного анализа (с использованием флюоресцентных зондов и меток).
Флюор.анализ – дает возможность исследовать подвижность фосфолипидных молекул в мембране, оценить вязкость липидной фазы.
|
№ 26 Электронная микроскопия, принцип устройства, предел разрешения электронного микроскопа. Рентгеноструктурный анализ, формула Вульфа - Брэггов. Какие характеристики биологических объектов изучают с помощью этих методов?
В электронном микроскопе вместо светового пучка на исследуемый объект направляется пучок электронов, разогнанных до больших скоростей. Известно, что электронам с высокими скоростями тоже присущи волновые свойства, в том числе явление дифракции. Однако при достаточно больших скоростях согласно формуле де Бройля, длина волны мала, и предел разрешения тоже мал.
В электроном микроскопе достигаются увеличения в сотни тысяч раз, что дало возможность исследовать строение клетки, клеточных органелл и биологических мембран.
Недостатки электронного микроскопа является деформация живого объекта в процессе исследования.
В электронном микроскопе носителем информации являются электроны, а источником – подогреваемый катод. Ускорение электрона и образование пучка осуществляется фокусирующим электродом и анодом – системой, называемой электронной пушкой. После взаимодействия с образцом поток электрона преобразуется и содержит информацию об образце. Формирование потока электронов происходит под действием электрического и магнитного полей.
Рентгеноструктурный анализ: позволяет обнаружить упорядоченность в расположении атомов и определять параметры упорядоченных структур (например, расстояние между кристаллографическими плоскостями) Исследование дифракции рентгеновских лучей на мембране подтвердили относительно упорядоченное расположение липидных молекул в мембране.
|
№ 27 Электрохимический потенциал, формула, физический смысл. Плотность потока вещества. Активный и пассивный перенос веществ через биологическую мембрану. Принципиальные различия между ними.
Э.Х.П. – работа, которую нужно совершить ( т.е. затратить энергию) чтобы перенести 1 моль вещества в электрическое поле вакуума и данной точки в бесконечно удаленную. r-численно равен энергии Гиббса, приходящийся на 1 моль вещества, помещенного в электрическом поле.
Д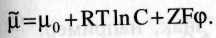 ля разбавленных растворов: ля разбавленных растворов:
где μ0 – химическая постоянная,К - газовая постоянная; Т — абсолютная температура; С – концентрация ионов; Z — заряд иона, F - число Фарадея — электрический заряд одного моля одновалентных ионов); φ — электрический потенциал.
Плотность потока вещества: 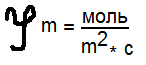 количество вещества, переносимого за единицу времени через единицу площади. Пассивный транспорт - это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением 1) идет самопроизвольно 2) Без затрата энергии АТФ. УРАВНЕНИЕ ТЕОРЕЛЛА: количество вещества, переносимого за единицу времени через единицу площади. Пассивный транспорт - это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением 1) идет самопроизвольно 2) Без затрата энергии АТФ. УРАВНЕНИЕ ТЕОРЕЛЛА: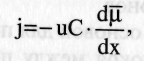 где j — плотность потока вещ-ва, u - подвижность частиц; С — концентрация; dμ / dx - градиент электрохимического потенциала, т.е. величина, показывающая скорость его изменения с расстоянием х. где j — плотность потока вещ-ва, u - подвижность частиц; С — концентрация; dμ / dx - градиент электрохимического потенциала, т.е. величина, показывающая скорость его изменения с расстоянием х.
УРАВНЕНИЕ НЕРНСТА – ПЛАНКА: 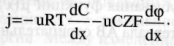 Активный транспорт - это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением. Из уравнения нернста планка следует что существует 2 причины переноса вещества: градиент электрохим. Потенциала и градиент концентрации. 1) Не может идти самопроизвольно. 2) Происходит за счет гидролиза АТФ. Активный транспорт - это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением. Из уравнения нернста планка следует что существует 2 причины переноса вещества: градиент электрохим. Потенциала и градиент концентрации. 1) Не может идти самопроизвольно. 2) Происходит за счет гидролиза АТФ.
|
|
|
№ 28 Пассивный транспорт заряженных частиц: Уравнение Теорелла, уравнение Нернста – Планка. Что такое градиент концентрации и градиент электрического потенциала?
Диффузия — самопроизвольное перемещение вещества из мест с большей концентрацией в места с меньшей концентрацией вещества вследствие хаотического теплового движения молекул.
Пассивный транспорт - это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением , без затрата энергии. УРАВНЕНИЕ ТЕОРЕЛЛА: где j — плотность потока вещ-ва, u - подвижность частиц; С — концентрация; dμ / dx - градиент электрохимического потенциала, т.е. величина, показывающая скорость его изменения с расстоянием х.
УРАВНЕНИЕ НЕРНСТА – ПЛАНКА:
Градиент: Знаки минусов перед градиентами показывают, что градиент концентрации вызывает перенос вещества от мест с большей конц. К местам с его меньшей конц. , а градиент электрического потенциала вызывает перенос положительных зарядов от мест с большим к местам с меньшим потенциалом.
|
№ 29 Пассивный транспорт не заряженных частиц: Закон Фика, осмос, фильтрация. Облегченная диффузия, ее свойства.
Диффузия — самопроизвольное перемещение вещества из мест с большей концентрацией в места с меньшей концентрацией вещества вследствие хаотического теплового движения молекул.
П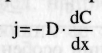 ассивный транспорт - это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением , без затрата энергии. ассивный транспорт - это перенос вещества из мест с большим значением электрохимического потенциала к местам с его меньшим значением , без затрата энергии.
ЗАКОН ФИКА: Уравнение описывающее простую диффузию.
где j — плотность потока вещ-ва, dС / dx - градиент концентрации, D –коэффициентДиффузии. Осмос – преимущественное движение молекул воды через полупроницаемые мембраны из мест с меньшей концентрацией растворенного вещества в места с большей концентрацией. Перенос растворителя (вода) из мест с меньшим количеством растворителя в места с большим. Фильтрация – движение раствора через поры в мембране под действием градиента давления.
Облегченная диффузия – перенос вещества с помощью молекул – переносчиков. Ее отличия от простой: 1) перенос вещества с участием переносчика происходит быстрее 2) обладает свойством насыщения: при увелич. Конц. С одной стороны мемб. Плотность потока вещ-ва возрастает, лишь до некоторого предела , когда все молекулы переносчика уже заняты. 3) При облег. Диф. Наблюдается КОНКУРЕНЦИЯ переносимых в-в в тех случаях, когда переносчиком переносятся разные в-ва.
|
№ 30 Активный транспорт веществ. Опыт Уссинга.
А.Т.В – это перенос вещества из мест с меньшим значением электрохимического потенциала в места с его большим значением. Активный транспорт в мембране сопровождается ростом энергии Гиббса, он не может идти самопроизвольно, а только в сопряжении с процессом гидролиза аденозинтрифосфорной кислоты (АТФ) т.е. за счет затраты энергии, запасенной в макроэргических связяк АТФ. Активный транспорт веществ через биологические мембраны впервые был доказан в опытах Уссинга (1949 г.) на примере переноса ионов натрия через кожу лягушки. Экспериментальная камера Уссинга, заполненная нормальным раствором Рингера, была разделена на две части свежеизолированной кожей лягушки. Наблюдались потоки ионов натрия через кожу: слева направо от наружной к внутренней поверхности jMвн и справа налево от внутренней к наружной поверхности — jM нар. Из уравнения Теорелла, описывающего активный транспорт, следует уравнение Уссинга-Теорелла для отношения этих потоков в случае пассивного транспорта:
г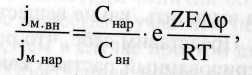 де Сна и Свн — концентрации натрия в наружном и внутреннем растворах; Δφ - возникшая разность потенциалов; F - постоянная Фарадея; R — универсальная газовая постоянная; Т — абсолютная температура; е = 2,71 (основание натурального логарифма). де Сна и Свн — концентрации натрия в наружном и внутреннем растворах; Δφ - возникшая разность потенциалов; F - постоянная Фарадея; R — универсальная газовая постоянная; Т — абсолютная температура; е = 2,71 (основание натурального логарифма).
На коже лягушки, разделявшей раствор Рингера, возникала разность потенциалов (Фвн -фнар). Внутренняя сторона кожи имела положительный потенциал по отношению к наружной.
В установке Уссинга имелся специальный блок компенсации напряжения, с помощью которого устанавливалась разность потенциалов на коже лягушки, равная нулю, что контролировалось вольтметром.
Кроме того, поддерживалась одинаковая концентрация ионов с наружной и внутренней стороны Снар=Свн, чем ликвидировались причины пассивного транспорта. При этих условиях, согласно уравнению Уссинга—Теорелла, потоки jM вн и jM нар должны быть равны друг другу: jM.BH=jmнар Естественно, что суммарный поток через мембрану был в этом случае равен нулю. Однако в условиях опыта даже при равенстве нулю градиентов электрического потенциала и концентрации через кожу лягушки протекал электрический ток, измеряемый амперметром. Из этого следовало, что происходил односторонний перенос заряженных частиц. Это неопровержимо свидетельствовало о том, что наряду с пассивным транспортом через кожу осуществлялся и активный перенос ионов .
|
|
|
№ 31 Что такое мембранная разность потенциалов? Схема регистрации мембранной разности потенциалов с помощью микроэлектрода. Формула Нернста для расчета равновесной мембранной разности потенциалов. Уравнение Гольдмана.
Одна из важнейших функций биологич.мембраны – генерация и передача биопотенциалов. Это явление лежит в основе возбудимости клеток, регуляции внутриклеточных процессов, работы нервной системы, регуляции мышечного сокращения.
В процессе жизнедеятельности в клетках и тканях могут возникать разности электрических потенциалов: 1) Окислительно-восстановительные потенциалы – вследствие переноса электронов от одних молекул к другим. 2) мембранные – вследствие градиента концентрации ионов и переноса ионов через мембрану.
Мембранный потенциал – называется разность потенциалов между внутренней и наружной поверхностями мембраны.
 Стелкянный микроэлектрод представляет собой стеклянную микропипетку с оттянутым очень тонким кончиком. Стелкянный микроэлектрод представляет собой стеклянную микропипетку с оттянутым очень тонким кончиком.
Металический электрод не может проколоть клеточную мембрану, кроме того он поляризуется. Для исключения поляризации электрода используются неполяризующиеся электроды, например серебряная проволка, покрытая солью AgCl.
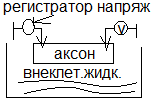
В раствор KCL или NACL, заполняющий микроэлектрод. Второй электрод – электрод сравнения – располагается в растворе в наружной поверхности клетки. Регистрир. Устройство Р, содержащее усилитель постоянного тока измеряет мембранный потенциал
Микроэлектродный метод дал возможность измерить биопотенциалы не только на гигантском аксоне кальмапа, но и на клетках нормал. Размеров: нервных волокнах других животных, клетках миокарда. Мембранные потенциалы подразделяются на потенциалы покоя и потенциалы действия. Формула НЕРНСТА: 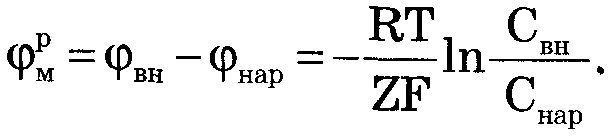 Уравнение Гольдмана: Уравнение Гольдмана:
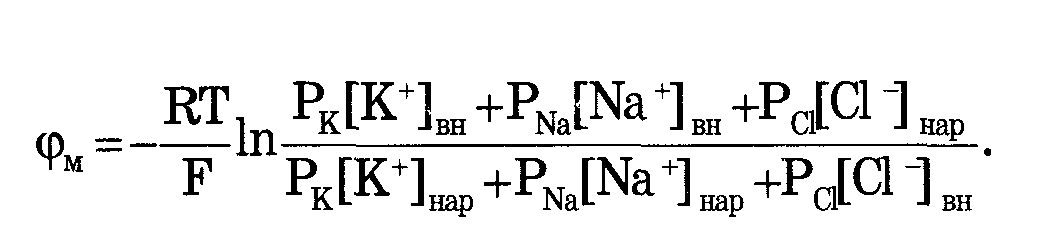 С Соотношением проницаемости PK : PNa : PCL = 1 : 0.05 : 0.45 С Соотношением проницаемости PK : PNa : PCL = 1 : 0.05 : 0.45
|
№ 32 Как возникает биопотенциал покоя? Напишите уравнение Нернста – Планка, объясните роль градиентов концентрации и электрического потенциала при формировании потенциала покоя.
Потенциал покоя - стационарная разность электрических потенциалов, регистрируемая между внутренней и наружной поверхностями мембраны в невозбужденном состоянии. Потенциал покоя определяется разной концентрацией ионов по Разные стороны мембраны и диффузией ионов через мембрану. Если концентрация какого-либо иона внутри клетки С отличив от концентрации этого иона снаружи С и мембрана проницала для этого иона, возникает поток заряженных частиц через Мембрану, вследствие чего нарушается электрическая нейтральность системы, образуется разность потенциалов внутри и снаружи клетки , которая будет препятствовать дальнейшему перемещению ионов через мембрану.
УРАВНЕНИЕ НЕРНСТА – ПЛАНКА: Градиент : Знаки минусов перед градиентами показывают, что градиент концентрации вызывает перенос вещества от мест с большей конц. К местам с его меньшей конц. , а градиент электрического потенциала вызывает перенос положительных зарядов от мест с большим к местам с меньшим потенциалом.
Причины формирования и стационарности:
1)Разные концентрации ионов внутри и снаружи мембраны
2)Диффузия
3)Различная проницаемость ионов
4)Насосы (активный транспорт)
|
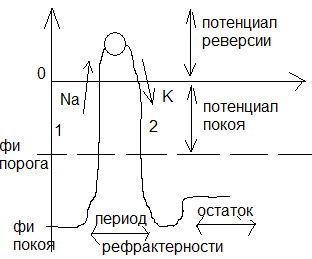
№ 33 Схема регистрации потенциала действия в аксоне. График потенциала действия. Относительные проницаемости мембраны для ионов К и Na в покое и при возбуждении.
 Потенциал действия – называется электрический импульс возникающий при внешнем стимуле на мембрану, обусловленный резким возрастанием проницаемости ионов Na. Соотношением проницаемости Потенциал действия – называется электрический импульс возникающий при внешнем стимуле на мембрану, обусловленный резким возрастанием проницаемости ионов Na. Соотношением проницаемости
PK : PNa : PCL = 1 : 0.05 : 0.45
Характерные свойства потенциала действия
1)Наличие порогового значения деполяризующего потенциала.
2)Закон все или ничего! Потенциал действия либо образуется либо нет
3) Наличие периоды рефрактерности (т.е. невозбудимости) С момента когда потенциал сдвинулся с потенциала покоя и до тез пор пока к этому значению не вернется мембрана невозбудима, т.к. никакой другой стимул отклика не дает.
4) Амплитуда потенциала действия не зависит от амплитуды внешнего стимула
5) В момент проведения потенциала действия проницаемость мембраны увеличивается а сопротивление уменьшается.
|
|
№ 34 Уравнение для тока мембраны при возбуждении (уравнение Ходжкина – Хаксли). Метод фиксации потенциала. Суммарный, калиевый и натриевый ионные токи в процессе возбуждения мембраны, их графики. Для чего используют блокаторы ионных потоков через мембрану?
Уравнение Хюджина – Хаксли описывает суммарные токи, обусловленные разными ионами
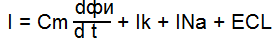
Емкостный ток обусловлен перезарядкой конденсатора, который представляет собой мембрана, перетеканием зарядов с одной ее поверхности на другую.
Каждый ионный ток определяется разностью мембр.потенциала , и равновесного нернстов потенц, создаваемого диффузией ионов данного типа.
Согласно теории Хюджина – Хаксли, возбуждении элемента мембраны связано с изменением проводимости мембраны для ионов натрия и кальция.
Метод фиксации. Была сконструирована специальная электрическая схема, которая позволяла фиксировать мембранный потенциал на определенном значении. Это позволило: 1) Избавиться от емкостных токов. 2) избавиться от зависимости g (фи) и излучать g/t)
Результаты:
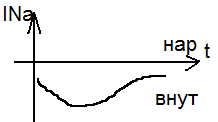
1) Введение специал. Вещества ТЭА (тетраэтиламмония), заблокировало К- каналы, и изучаем ток Na.
2) Заблокировали Na-каналы(ТТХ – тетродоксин, изучаем ток К)
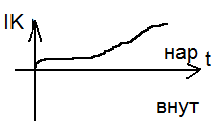
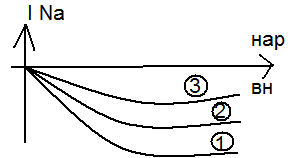 Чем дальше ступенька от потенциала покоя, тем чегото натриевый ток. Чем дальше ступенька от потенциала покоя, тем чегото натриевый ток.
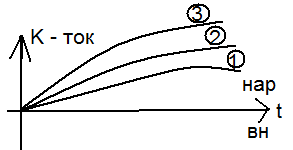 Чем дальше от покоя тем выше ток К. Чем дальше от покоя тем выше ток К.
Проводимость натриевого и калиевого каналов зависит от времени и величины мембранного потенциала.
Селективное изменение ионной проницаемости возбужденной мембраны сначала для Na +, потом для К+ объясняется тем, что в мембране имеются специальные ионные каналы. (имеют белковое происхождение)
Одним из доказательств наличия в мембране ионных каналов
|
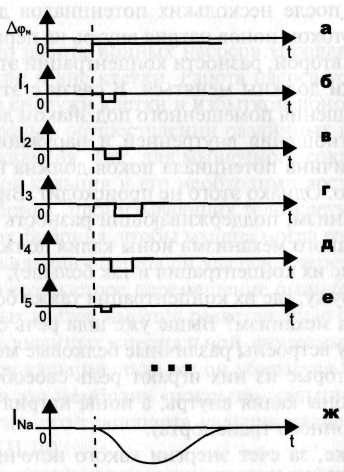
№ 35 Токи одиночных натриевых каналов. Структура натриевых каналов, их свойства: дискретность и взаимно независимость действия, селективность, время открытого состояния канала.
Канал за время одного деполяризующего сдвига открывался лишь один раз на время tи, которое будем называть временем открытого состояния канала.
Среднее значение tи для Nа+ канала = 0,7 мс (от 0,3 до 1,5 мс). Одиночный канал может открыться раньше или позже.
Время, в течение которого вероятность открывания отдельного канала велика, будем называть временем жизни каналов: ТNa, ТCa. Для натриевых каналов ТNa= 2 мс.
Свойства ионных каналов: 1)Селективностью называют способность ионных каналов избирательно пропускать ионы какого либо одного типа.
2) Независимость работы отдельных каналов – ток через тот или иной канал не зависит от того протекает ли ток через другой и канал или нет. 3) Дискретный характер проводимости ионных каналов. – ионный канал может находится в 2х состояниях: открытом или закрытом. Переходы между состояниями, происходят в случайные моменты.
4) Зависимость параметров канала от мембранного потенциала.(при изменении потенциала меняются токи)
Состояние натриевого канала – 1) ПОКОЙ (ворота закрыты т.к. потенциал меньше потенциала порога) 2) АКТИВАЦИЯ (вероятность открытого состояния резко возрастает, т.к. больше потенциала порога
3) САМОПРАИЗВОЛЬНАЯ ИНАКТИВАЦИЯ (вероятность открыть состоящ. Убывает, несмотря на то что потенциал больше потенциала порога
4) ПОКОЙ (закрыт т.к. снова потенциал меньше потенциала порога)
|
№ 36 Механизм распространения потенциалов действия вдоль нервного волокна, локальные токи. Почему по неразветвленному аксону возбуждение распространяется только в одну сторону?
Если в каком нибудь участке возбудимой мембраны сформировался потенциал действия, мембрана деполяризована, возбуждение распространяется на др. участки мембраны.
Возникают локальные токи и внутри аксона, и на наружной поверхности (между участками поверхности мембраны с большим потенциалом + и участками с меньшим потенциалом - .
Локальные электрические токи приводят к повышению внутреннего потенциала поверхности невозбужденного участка мембраны вн потенциала и к понижению наружного потенциала невозбужденного участка мембраны, оказавшегося по соседству с возбужденной зоной.
Мембранный потенциал возрастает под действием изменений мемб.потенциала открываются натриевые каналы.
Происходит деполяризация мембраны, развивает потенциал действия. Затем возбуждение передается дальше на покоящиеся участки мембраны.
Возбуждение может распространяться только в область мембраны, находящуюся в состоянии покоя, т.е. в одну сторону от возбужденного участка аксона.
|
|
|
№ 37 Соотношение ионов Na+, Ca+ и K+ внутри и снаружи кардиомиоцита. Потенциал действия кардиомиоцита, график. Состояния каналов и направления потоков ионов Na+, Ca+ и K+ в различные фазы потенциала действия кардиомиоцита.
Ионные насосы миокардиальных клеток. Сохранение ионного баланса в кардиомиоцитах обеспечивает К+- Na+ и Са2+-насосы, активно перекачивающие ионы Na+ и Са+ наружу, а ионы K+ - внутрь клетки Работу этих насосов обеспечивают ферменты K+- Na+ АТФаза и Са2+-АТФаза, находящиеся в сарколемме миокардиальных клеток. Потенциал действия клетки миокарда имеет три характерные фазы: деполяризация (I), плато (II) и реполяризация (III).
I фаза — деполяризация, определяется резким ростом проницаемости мембраны для ионов натрия:
Рк : PNa = 1 : 20 в момент превышения фим порогового значения при возбуждении. Порог активации натриевых каналов примерно -60 мВ, а время жизни 1 - 2 мс и может доходить до 6 мс.
I фаза — плато - характерна медленным спадом фим от пикового значения (= + 30 мВ) до нуля. В этой фазе одновременно работают два типа каналов - медленные кальциевые каналы и калиевые каналы. Кальциевые каналы имеют порог активации около -30 мВ, а время их жизни примерно 200 мс. В результате открывания кальциевых каналов возникает деполяризующий медленный входящий в клетку кальциевый ток: Одновременно с ростом кальциевого тока растет проводимость для ионов калия, что приводит к возникновению вытекающего калиевого тока, реполяризующего мембрану. Во II фазе gp^ уменьшается, a g^ увеличивается (см. рис. 4.9), происходит постепенное выравнивание текущих навстречу друг Другу токов, а потенциал мембраны (р понижается почти до нуля. Для II фазы характерно, что суммарный ток мембраны I стремится к 0, то есть II фаза — реполяризация - характеризуется закрытием кальциевых каналов, ростом величины gк и усилением выходящего тока K+.
Модифицируя уравнение (3.8), можно получить уравнение для мембранного тока при возбуждении кардиомиоцита:
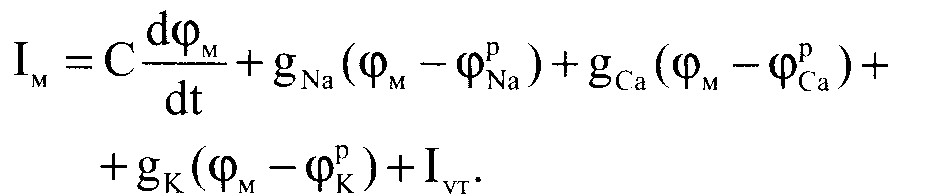
|
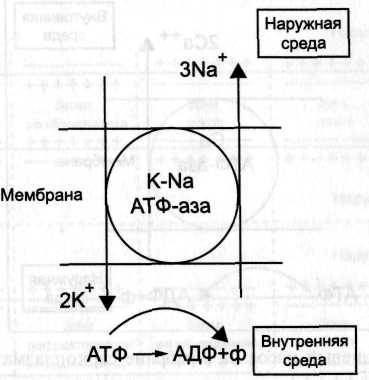
№ 38 Какую функцию выполняют ионные насосы в мембранах? Схема действия K–Na–насоса. Сопряженные процессы в ионных насосах.
Ионные насосы миокардиальных клеток. Сохранение ионного баланса в кардиомиоцитах обеспечивает К+- Na+ и Са2+-насосы, активно перекачивающие ионы Na+ и Са+ наружу, а ионы K+ - внутрь клетки Работу этих насосов обеспечивают ферменты K+- Na+ АТФаза и Са2+-АТФаза, находящиеся в сарколемме миокардиальных клеток.
для закачивания внутрь клетки 2 ионов К и выкачивания наружу 3х ионов Na достаточно энергии распада одной молекулы АТФ.
- 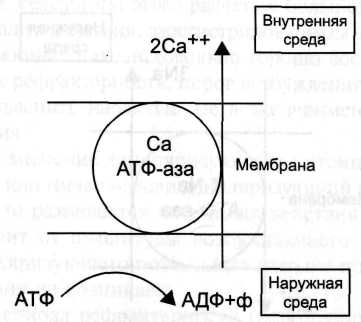 для мышечного сокращения необходимо много ионов Са и его необходимо доставлять в каждой из белковых фибрилл. Внутри мышечных клеток имеется разветвленная система полостей и трубочек, образов. Специальной внутр. Мембраной, в которых хранится Са. Вся эта мембрана покрыта кальциевыми насосами. для мышечного сокращения необходимо много ионов Са и его необходимо доставлять в каждой из белковых фибрилл. Внутри мышечных клеток имеется разветвленная система полостей и трубочек, образов. Специальной внутр. Мембраной, в которых хранится Са. Вся эта мембрана покрыта кальциевыми насосами.
|
№ 39 Как регистрируют электрокардиограмму? Основные положения теории Эйнтховена. Объясните Генез ЭКГ на базе дипольной модели.
Метод исследования работы органов и тканей, основанный на регистрации во времени потенциалов электр. Поля на поверх. Тела называется электрографией. Два электрода, приложенные к разным точкам на поверхности телаЖ, регистрируют разность потенциалов. Временная зависимость изменения этой разности потенциалов называется ЭЛЕКТРОГРАММОЙ. (Экг – сердца, ЭРГ – сетчатки глаза, ЭЭГ – головного мозга, ЭМГ – мышц.)
ЭЛЕКТРОКАРДИОГРАММА – ЭКГ – это график временной зависимости разности потенциалов в соответствующие отведения, а значит и временной зависимости проекции ИЭВС на линию отведения.
ИЭВС – интегр. Электр.вектор сердца.
Гармонический спектр ЭКГ – в основном содержит частоты от 0 до 100 Гц.
Теория ЭЙНТХОВЕНА. Эйнтховен предложил измерять разности потенциалов между двумя из 3х точек , представл. Вершины треугольника.
1 отведение – м\д Левой рукой и п. рукой.
2 отведение – м\д л.ногой и прав. рукой
3 отведение – м\д л.ногой и лев рукой
Руки и ноги рассматривались как проводники отводящие потенциалы от времени треугольника Эйнтховена.
1)Сердце можно представить как эквивалентый генератор , имеющий дипольную природу
2) каждая клетка сократительного миокарда обладает дипольным маментом
3) ИЭВС –находится в однородной проводящей среде – начало неподвижно, находится в атриовентрикулярном усле, в конец описывают кривую из 3х петель.
4) суммар.дипольный момент сердца.
|
|
|
№ 40 Электроэнцефалография. Использование теоремы Фурье для спектрального анализа ЭЭГ. Метод картирование электрической активности мозга при анализе ЭЭГ.
Электроэнцефалография применяется в медицине для определения области опухоли мозга, для оценки функционального состояния мозга до и после введения лекарственного препарата.
Электроэнцефалограмма - это график изменения разности потенциалов между различными участками (точками съема) поверхности головы человека во времени. Количество точек съема может существенно меняться (от 2 до нескольких десятков) в зависимости от целей исследования. ЭЭГ отражает интегральную активность огромного числа нейронов коры головного мозга и распространение волн возбуждения в нейронных сетях. Электроэнцефалограмма имеет вид сложных регулярных колебаний с различными частотами и амплитудой. Для исследования электрической активности мозга при различных функциональных состояниях обычно рассматриваются спектральные составляющие (простые синусоидальные колебания различных частот и амплитуд, на которые, согласно теореме Фурье, можно разложить сложное колебание - электроэнцефалограмму).
Далее решаются задачи двух типов:
1. Находят полную мощность сигнала ЭЭГ в каждом отведении (площадь под соответствующей спектральной кривой) и строят карту распределения полной мощности по поверхности головы, присваивая каждому из интервалов, на которые разбит весь промежуток значений мощности, свой цвет или оттенок. 2. Находят не полную мощность сигнала в том или ином отведении, а лишь мощность, соответствующую тому или иному спектральному интервалу.
|
№ 41 Что такое среды (АС), каковы их свойства. Автоволны, особенности распространения автоволн в АС. Тау-модель, длина волны возбуждения. Примеры АС и автоволн в организме.
Активная среда (AC) - это среда, состоящая из большого числа отдельных элементов (например, клеток), каждый из которых является автономным источником энергии. Элементы активной среды имеют контакт между собой и могут передавать импульс возбуждения от одной клетки к другой. Примером активной среды в организме являются нервные волокна и нейронные сети, мышечные структуры сердца, гладкомышечные волокна сосудов, желудка, а также другие ткани. В таких средах распространяются волны возбуждения, называемые автоволнами. Математическое описание процессов распространения автоволн в активных средах связано с решением систем уравнений. Решение этих систем представляет значительные трудности. Поэтому для описания автоволновых процессов используются модели формальных активных сред, например, модель, предложенная Винером и Розенблютом, называемая тау-моделью. В этой модели постулируется, что каждая клетка, являющаяся элементом активной среды, может находиться в одном из трех состояний: 1) возбуждение - тау, если ее φм > φмпор ; в этом состоянии клетка не возбудима, но может возбудить соседнюю клетку, находящуюся в покое; 2) "рефракторный" хвост - (R-τ), если φмп < φм < φмпор; в этом состоянии эта клетка не возбудима, но не может возбудить клетку, находящуюся в покое; 3) покой - ее φм = φмп ; в этом состоянии клетка может быть возбуждена соседней при условии, что трансмембранный потенциал соседней клетки выше значения порога рассматриваемой.
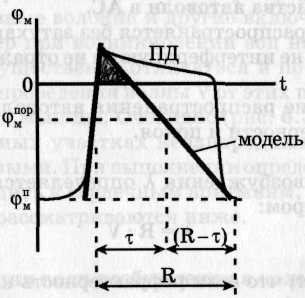 Автоволны - это самоподдерживающиеся волны возбуждения в активной среде, сохраняющие свои характеристики постоянными за счет распределенных в среде источников энергии. Характеристики волны - период, длина волны, скорость распространения, амплитуда и форма - в установившемся в режиме зависят только от локальных свойств активной среды и не зависят от начальных условий. Автоволны - это самоподдерживающиеся волны возбуждения в активной среде, сохраняющие свои характеристики постоянными за счет распределенных в среде источников энергии. Характеристики волны - период, длина волны, скорость распространения, амплитуда и форма - в установившемся в режиме зависят только от локальных свойств активной среды и не зависят от начальных условий.
|
№ 42 Распространение автоволн в неоднородных по рефрактерности активных средах. Трансформация ритма, условия её возникновения.
Неоднородной называется активная среда, в различных участках которой значения R и V могут быть не одинаковыми. Активная среда организма, например мышечная ткань, неоднородна. В разных участках мышцы могут проходить кровеносные сосуды, нервные волокна и другие включения. При патологиях, например при возникновении зон некроза, свойства этих зон могут существенно отличаться и по рефрактерности R, и по скорости проведения волны V от этих параметров в участках нормальной мышцы. Очевидно, что длины автоволн в различных участках неоднородных активных сред будут неодинаковыми.
В проводящий системе сердца, а также в самой сердечной мышце могут образовываться замкнутые пути, по которым циркулирует волна возбуждения.
Если период рефрактерности участка среды R2 больше периода рефрактерности остальной части среды R1 и если интервал между посылкой двух импульсов возбуждения Т меньше периода рефрактерности R2: Т < R2, вторая волна не может возбудить область с R2 > R1.
|
|
| |
|
|
 Скачать 5.18 Mb.
Скачать 5.18 Mb.