1. Вирусология (virus яд животного происхождения, logos наука)
 Скачать 0.66 Mb. Скачать 0.66 Mb.
|

  1. Место вирусологии в ряду классических наук. Содержание фундаментальных и прикладных разделов вирусологии, общей и частной вирусологии. Связь вирусологии с другими областями знаний. 1. Место вирусологии в ряду классических наук. Содержание фундаментальных и прикладных разделов вирусологии, общей и частной вирусологии. Связь вирусологии с другими областями знаний.1. Вирусология (virus – яд животного происхождения, logos - наука) – это биологическая наука, занимающаяся изучением мельчайших микроорганизмов - вирусов Она изучает природу, морфологию, химический состав, взаимоотношения с организмом и внешней средой, методы диагностики, профилактики и лечения вирусных болезней. Вирусология делится на: Общую: включает в себя общую характеристику и систематику вирусов; взаимодействие вирусов с клетками; культивирование, экологию и генетику вирусов; патогенез вирусных инфекций; особенности противовирусного иммунитета; принципы лабораторной диагностики, лечения и специфической профилактики вирусных болезней. Частную: изучает те же вопросы, но применительно к конкретному возбудителю. Связь вирусологии с другими науками и ее задачи. Вирусология – молодая наука. Она тесно связана с другими науками и прежде всего с микробиологией. Вирусологи широко используют бактериологические методы: фильтрация, стерилизация, микроскопия, серологические реакции. При изучении структуры вирусов, взаимодействия их с клетками используются методы цитологии. Очень тесно связана вирусология с биохимией, с генетикой. Вирусология дает необходимые знания при изучении эпизоотологии, болезней мелких животных Одна из важнейших задач, стоящих перед вирусологией, - это изучение структуры, химического состава, биологии, генетики и селекции вирусо, взаимодействия вируса и клетки, устойчивости вирусов к разным факторам. Только успешное решение теоретических вопросов позволит разрабатывать эффективные методы диагностики и борьбы с вирусными инфекциями. 2. Природа вирусов, их отличие от известных живых организмов. Какая форма биотических связей (взаимоотношений) характерна для вирусов. Итак, вирусы состоят из нескольких компонентов: сердцевина - генетический материал (ДНК или РНК). Генетический аппарат вируса несет информацию о нескольких типах белков, которые необходимы для образования нового вируса. белковая оболочка, которую называют капсидом. Оболочка часто построена из идентичных повторяющихся субъединиц - капсомеров. Дополнительная липопротеидная оболочка. Она образована из плазматической мембраны клетки-хозяина. Она встречается только у сравнительно больших вирусов (грипп, герпес). Каждый компонент вирионов имеет определённые функции: белковая оболочка защищает их от неблагоприятных воздействий, нуклеиновая кислота отвечает за наследственные и инфекционные свойства и играет ведущую роль в изменчивости вирусов, а ферменты участвуют в их размножении. Более сложные по структуре вирусы, кроме белков и нуклеиновых кислот, содержат углеводы, липиды. Для каждой группы вирусов характерен свой набор белков, жиров, углеводов и нуклеиновых кислот. Некоторые вирусы содержат в своём составе ферменты. В отличие от обычных живых клеток вирусы не употребляют пищи и не вырабатывают энергии. Они не способны размножаются без участия живой клетки. Вирус начинает размножаться лишь после того, как он проникнет в клетку определенного типа. качественное отличие вирусов от про- и эукариот. 1) наличие одного типа нуклеиновой кислоты (ДНК или РНК); 2) отсутствие клеточного строения и белоксинтезирующих систем; 3) возможность интеграции в клеточный геном и синхронной с ним репликации. Паразитизм (от греч. parasitos – нахлебник, дармоед) – межвидовые взаимоотношения, при которых один вид (паразит) использует другой (хозяина) как среду жизни и как источник пищи. Паразитизм возник в процессе тесного контакта разных видов организмов на основе пищевых и пространственных связей и встречается на всех уровнях организации живого, начиная с вирусов и бактерий и кончая высшими растениями и многоклеточными животными. Паразитизм является одной из форм антагонистического симбиоза, совместного проживания разных видов организмов. например вирус табачной мозаики, вирус кори. Саттелиты: Вирусы-сателлиты представляют собой крайнюю форму паразитизма. Они относятся к вирусам, паразитирующим на генных продуктах, образованных другими, часто неродственными им вирусами. Один из простейших примеров — сателлит вируса некроза табака (STNV). Он полностью зависит в своей репликации от одновременного заражения клеток табака его инфекционным вирусом-помощником, вирусом некроза табака. 3. Архитектура вирионов. Типы симметрии. Номенклатура терминов, используемых для описания архитектуры вирионов. Внеклеточная форма вируса — вирион, предназначенная для сохранения и переноса нуклеиновой кислоты вируса. Под архитектурой вирионов понимают ультратонкую структурную организацию этих надмолекулярных образований, различающихся размерами, формой и сложностью строения. Для описания архитектуры вирусных структур разработана номенклатура терминов: Белковая субъединица — единая, уложенная определенным образом полипептидная цепь. Структурная единица (структурный элемент) — белковый ансамбль более высокого порядка, образованный несколькими химически связанными идентичными или неидентичными субъединицами. Морфологическая единица — группа выступов (кластер) на поверхности капсида, видимая в электронном микроскопе. Капсид — внешний белковый чехол или футляр, образующий замкнутую сферу вокруг геномной нуклеиновой кислоты. Кор (core) — внутренняя белковая оболочка, непосредственно примыкающая к нуклеиновой кислоте. Нуклеокапсид — комплекс белка с нуклеиновой кислотой, представляющий собой упакованную форму генома. Суперкапсид или пеплос — оболочка вириона, образованная липидной мембраной клеточного происхождения и вирусными белками. Матрикс — белковый компонент, локализованный между суперкапсидом и капсидом. Пепломеры и шипы — поверхностные выступы суперкапсида. 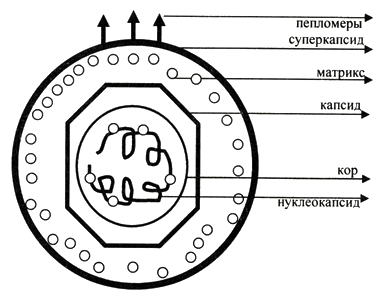 Рис. 1. Строение гипотетического вириона Рис. 1. Строение гипотетического вирионаВирионам или их компонентам могут быть присущи два основных типа симметрии (свойство тел повторять свои части) — спиральный и икосаэдрический. В том случае, если компоненты вириона обладают разной симметрией, то говорят о комбинированном типе симметрии ВЧ. (схема 1). Спиральная укладка. Классическим примером вируса со спиральным типом симметрии является вирус табачной мозаики (ВТМ). Нуклеокапсид этого палочковидного вируса размером 18x300 нм состоит из 2130 идентичных субъединиц, на виток спирали приходится 16 1/3 субъединиц, шаг спирали составляет 2,3 нм. Икосаэдрическая симметрия — самая эффективная для конструирования замкнутого чехла из отдельных субъединиц. При рассмотрении элементов икосаэдрической симметрии следует различать понятия симметрия и форма. Симметрия в данном случае — это набор поворотов, которые переводят объект сам в себя, форма — это лишь общий вид кубической поверхности объекта (тетраэдр, октаэдр, додекаэдр и т. д.). Икосаэдр — это геометрическая фигура, имеющая 12 вершин, 20 граней, 20 ребер. 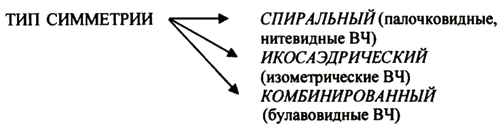 Еще более сложноустроенные вирионы, на пример частицы бактериофагов T-чётной серии, обладают комбинированным типом симметрии. Так, головка бактериофага T4 имеет икосаэдрический тип симметрии, а сокращенный чехол хвостового отростка обладает спиральным типом симметрии. 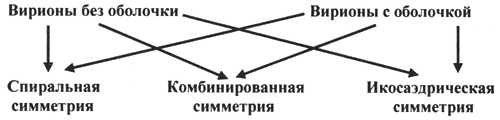 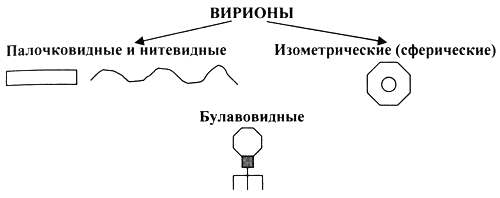 Рис. 2. Морфологические типы непокрытых вирионов 4. Морфологические типы вирусов. Особенности этих типов (связь белка с НК, соотношение белок/НК, поверхность взаимодействия с внешней средой). Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса).По форме вирионы могут быть: округлыми, палочковидными, в виде правильных многоугольников, нитевидными и др. Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы. По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму. Нуклеиновые кислоты вирусов.Вирусы содержат только один тип нуклеиновой кислоты, ДНК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). В состав простых вирионов входит один тип нуклеиновой кис лоты — РНК или ДНК — и белки. У сложных вирионов в составе внешней оболочки содержатся липиды и полисахариды, которые они получают из клеток хозяина. Вирусные ДНК. Молекулярная масса ДНК разных вирусов пример но в 10—100 раз меньше молекулярной массы ДНК бактерий. В геноме вирусов содержится до нескольких сотен генов. По сво ей структуре вирусные ДНК характеризуются рядом особен ностей, что дает возможность подразделить их на несколько типов. К ним относятся двунитевые и однонитевые ДНК, кото рые могут иметь линейную или кольцевую форму. Хотя в каж дой нити ДНК нуклеотидные последовательности встречаются однократно, на ее концах имеются прямые или инвертирован ные (повернутые на 180°) повторы. ДНК может быть: 1) двухцепочечной; 2) одноцепочечной; 3) кольцевой; 4) двухцепочечной, но с одной более короткой цепью;5) двухцепочечной, но с одной непрерывной, а с другой фрагментированной цепями. Вирусная РНК. Вирусные РНК по своему химическому составу не отличаются от РНК клеточного происхождения, но характеризуются разной структурой. Наряду с типичной для всех РНК однонитевой формой у ряда виру сов имеется двунитевая РНК. При этом она может быть линей ной и кольцевой. В составе однонитевых РНК имеются спираль ные участки типа двойной спирали ДНК, образующиеся вслед ствие спаривания комплементарных азотистых оснований. Одно нитевые РНК в зависимости от выполняемых ими функций подразделяют на две группы. К первой относят РНК, облада ющие способностью транслировать закодированную в ней инфор мацию на рибосомы клетки хозяина, т. е. выполнять функцию «РНК. Ее называют плюс-нить и обозначают знаком «+» (пози тивный геном). Ко второй группе относят вирусные одноцепочечные РНК, которые не могут функционировать как «РНК, а так же как ДНК служат лишь матрицей для ее образования. Такие РНК называют минус-нить, обозначают знаком «—» (не гативный геном). РНК может быть: 1) однонитевой; 2) линейной двухнитевой; 3) линейной фрагментированной; 4) кольцевой; 5) содержащей две одинаковые однонитевые РНК. Вирусные белки: структурные и функциональные. Первые входят главным образом в состав вирусного капсида, вторые представляют собой ферменты, участвующие в процессе репро дукции вирусов. Структурные белки у простых вирионов, лишенных суперкапсида, представлены капсидными белками, которые образуют фут ляр, защищающий нуклеиновую кислоту. У многих сложных вирионов в составе капсидных белков содержатся ферменты, участвующие в репликации и транскрипции вирусных РНК или ДНК. Кроме того, в составе вирионов имеются так называемые «внутренние» гистоноподобные белки, связанные с вирусной нуклеиновой кислотой. Они образуют рибо- или дезоксирибо-нуклеопротеиды, которые обладают определенными антигенными свойствами. 5. Белки вирусов, их происхождение и функции. Белки, связанные с жизненным циклом вируса, разделяют на белки, детерминируемые геномом вируса и белки, имеющие клеточное происхождение. 1)Клеточные белки: -белок цитоскелета — актин, и ядерные белки — гистоны. Белки клеточного происхождения,так же участвуют в процессе репликации вируса. По месту локализации белки, детерминируемые вирусным геномом, разделяют на две группы: 1) структурные белки — это белки, входящие в состав ВЧ, их обозначают как VP; 2) неструктурные белки — это предшественники структурных белков, регуляторные белки и ферменты, обслуживающие процесс внутриклеточной репродукции вируса и не входящие в состав ВЧ. Их обозначают как NS-белки . 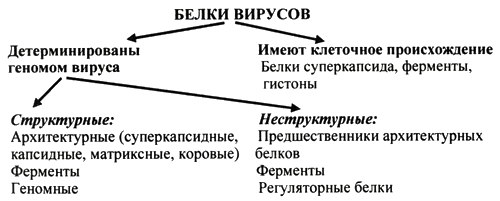 6. Функции гликопротеидов и липопротеидов вирусов. Происхождение вирусных липидов. Сложные белки представлены гликопротеинами и липопротеинами. Наличие гликопротеина определяет присутствие в вирионе углеводного компонента. Вирусные гликопротеины, как правило, экспонированы на наружной поверхности ВЧ и выполняют три основные функции: 1)обеспечивают связывание вириона с клеточным рецептором (функция прикрепительного белка), 2)обладают фузионной активностью (обеспечивают слияние мембран) 3) определяют антигенные свойства вирусов. В то же время, вирусные гликопротеины могут быть и неструктурными белками и, оставаясь в интегральной форме в мембране шероховатого эндоплазматического ретикулюма (ШЭР), выполнять функции транслоказ, обеспечивая транспорт вирусных компонентов в его просвет. Вирусные липопротеины представлены белками, ацилированными, как правило, миристиновой кислотой. Остатки жирных кислот, соединенные с молекулой белка, выполняют функцию липофильного якоря. 7. Какие нуклеиновые кислоты входят в состав вирусов и какие функции они выполняют. В отличие от клеток, вирионы содержат только один вид нуклеиновой кислоты — ДНК или РНК. И та и другая являются хранителями наследственной информации и выполняют функции генома. Однако следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса. В жизненном цикле вируса его геномная нуклеиновая кислота транскрибируется, то есть ДНК-содержащие вирусы образуют РНК. Ряд РНК-содержащих вирусов имеют в цикле репродукции стадию обратной транскрипции и синтезируют ДНК на матрице РНК. Примерно 20% всех вирусов имеют ДНК-геном, 80% — РНК-геном. Способность РНК хранить наследственную информацию — уникальное свойство вирусов. Размеры вирусных геномов (длина нуклеотидных последовательностей, выраженная в нуклеотидах) варьируют в широких пределах — от 1,7 тысяч нуклеотидов (т.н.) у цирковируса свиней до 300 т.н. у фикоднавирусов архибактерий. Кроме того, что геном вирусов может быть представлен или ДНК или РНК, он может находиться в разных видах — в виде двухнитевой (дн) или однонитевой (он) формы, в виде линейной или кольцевой, в виде непрерывной или сегментированной формы 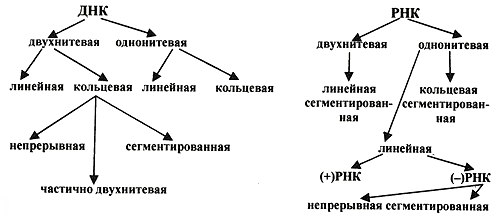 Многообразие видов РНК геномов расширяется за счет существования последовательностей, отличающихся направлением связей сахаро-фосфатного остова. Многообразие видов РНК геномов расширяется за счет существования последовательностей, отличающихся направлением связей сахаро-фосфатного остова.8. Вид (форма) ДНК-геномов вирусов. ДНК-содержащие вирусы – вирусы, геном которых представлен дезоксирибонуклеиновой кислотой и репликация идет посредством ДНК-зависимой ДНК-полимеразы, без использования промежуточного звена-посредника РНК. У многих ДНК-содержащих вирусов транскрипция происходит в три этапа, во время которых избирательно считываются сверхранние, ранние и поздние гены. Вирусы классифицируются 1)на те, которые содержат ДНК (вирус простого герпеса ) и те, что содержат РНК ( вирус иммунодефицита человека ). 2)По структуре капсомеров. Изометрические (кубические), спиральные, смешанные. 3)По наличию или отсутствию дополнительной липопротеидной оболочки 4)За клетками-хозяевами Геномная ДНК этих вирусов может быть двуцепочечной или одноцепочечной и иметь линейную или кольцевую Форму. 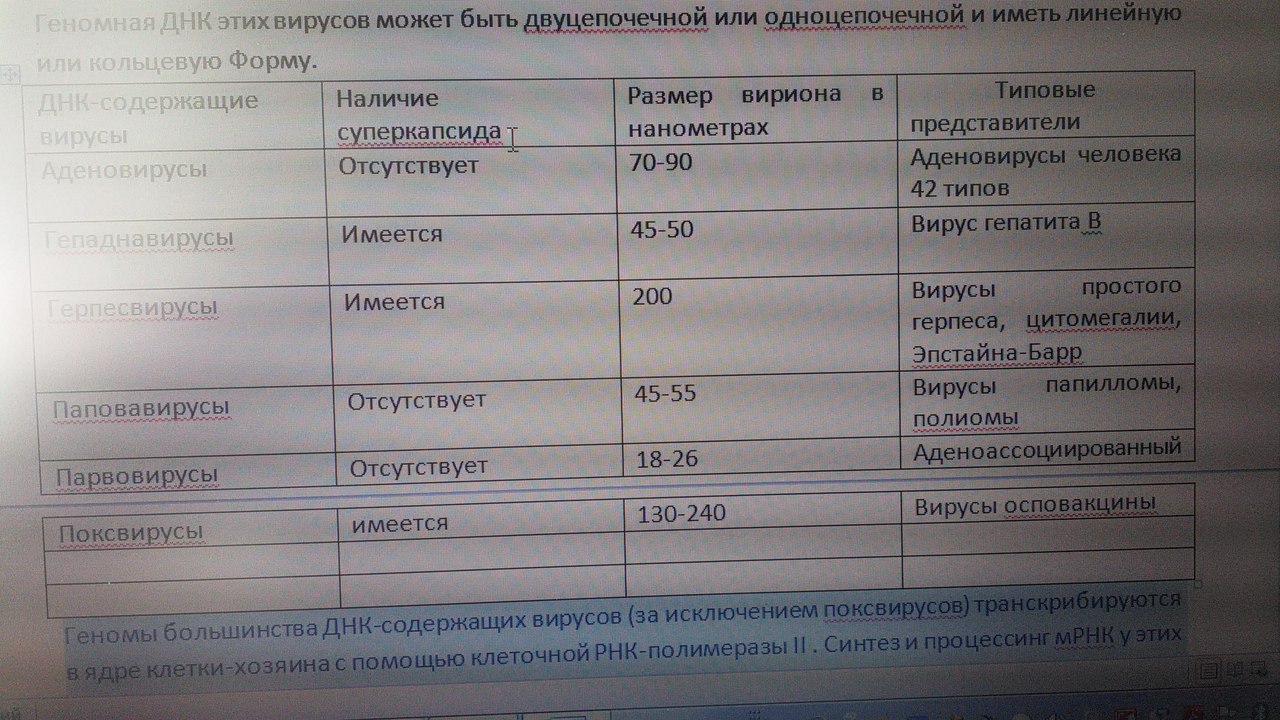 Геномы большинства ДНК-содержащих вирусов (за исключением поксвирусов) транскрибируются в ядре клетки-хозяина с помощью клеточной РНК-полимеразы II . Синтез и процессинг мРНК у этих вирусов практически полностью осуществляется за счет клеточных механизмов (несколько вирусных белков участвуют только в регуляции транскрипции). У многих ДНК-содержащих вирусов транскрипция происходит в три этапа, во время которых избирательно считываются сверхранние, ранние и поздние гены. 9. Вид (форма) РНК-геномов вирусов. Структура вирусных РНК чрезвычайно разнообразна. У вирусов обнаружены типы РНК: 1.Линейная односпиральная. Вирусы, содержащие однонитчатые РНК, делятся на две группы. У вирусов первой группы вирусный геном обладает функциями информационной РНК, т. е. может непосредственно переносить закодированную в нем информацию на рибосомы (обозначены знаком «плюс» и в связи с этим вирусы, содержащие такие РНК (пикорнавирусы, тогавирусы, коро-навирусы, ретровирусы), обозначены как «плюс-нитевые» вирусы, или вирусы с позитивным геномом.) Вторая группа РНК-содержащих вирусов содержит геном в виде однонитчатой РНК, которая сама не обладает функциями иРНК. В этом случае функцию иРНК выполняет РНК, комплементарная геному. Синтез этой РНК (транскрипция) осуществляется в зараженной клетке на матрице геномной РНК с помощью вирусспецифиче-ского фермента — транскриптазы (обозначаются как «минуса-РНК, а вирусы этой группы как «минус-нитевые» вирусы, или вирусы с негативным геномом.) К этим вирусам относятся ортомиксовирусы, парамиксовирусы, буньявирусы, рабдовирусы. 2.Линейная фрагментированная. 3.Кольцевая односпиральная. РНК-фрагменты буньявирусов обнаружены в виде кольцевой формы. 4.Линейная двуспиральная фрагментированная. Диплорнавирусыимеют двунитевое фрагментированное состояние генома. 5.Обоюдозначащая РНК Существуют вирусы, содержащие как «плюс-нитевые», так и «минус-нитевые» РНК гены (амбисенс-вирусы). К ним относятся аренавирусы. 10. Какие вещества кроме белка и НК входят в состав вирусов. Их функции. Помимо белков в липопротеидной оболочки обнаружены липиды и углеводы. Углеводы преимущественно содержаться в гликопротеидных пепломерах на поверхности вирусной частицы.Выросты, состоящие из липо–или гликопротеидов, выполняют рецепторную функцию. Т.е. у вирионов сложноорганизованных вирусов имеется еще поверхностная оболочка - суперкапсид, содержащий, кроме белков, также углеводы, липиды, компоненты клетки хозяина.Эти рецепторы впоследствии помогают вирусу присоединиться к клетке хозяина и потом проникнуть в нее. А липопротеидная оболочка вируса сливается с мембраной клетки, тем самым обеспечивая прохождения НК вируса в клетку. В составе вирусов обнаружены минеральные вещества К, Na, Ca, Mg, Fe. Они участвуют в формировании связей белка с нуклеиновой кислотой. 11. Что такое рецепторы и антирецепторы. Их состав. Их функция и на каком принципе она основана. Антирецепторы вирусные - поверхностные вирионные белки, напр., гемагглютинин, связывающиеся по комплементарному типу с соответствующим рецептором восприимчивой клетки. Первая фаза репликации вируса.Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды.Структура рецепторов - это их организация, принцип которой почти у всех рецепторов одинаков. Структурно рецептор состоит из трех доменов (частей): 1) внемембранного, осуществляющего взаимодействие с веществом (лигандом); 2) трансмембранного, осуществляющего перенос сигнала; 3) цитоплазматического. Эти рецепторы впоследствии помогают вирусу присоединиться к клетке хозяина и проникнуть в нее. 12. Способы проникновения вирусов в клетку. Механизм реализации проникновения. На настоящий момент термином "вход" обозначается четыре этапа инициации жизненного цикла вирусов в клетках-мишенях. К первому этапу относится присоединение вирионов к рецепторам клеточной мембраны, при котором происходят первоначальные конформационные изменения белков наружной оболочки вирусов. Ко второму, - взаимодействие с корецепторами посредниками проникновения вируса в клетку. Этот процесс сопровождается дальнейшими конформационными изменениями белковых компонентов вирусной оболочки, а также изменениями клеточной плазмалеммы. Третьим этапом является непосредственное перемещение вируса в клеточной мембране, обозначаемое термином проникновение или "penetration" и осуществляемое с помощью различных механизмов. К заключительному - четвертому, этапу входа относится освобождение генома вируса от нуклеокапсидной оболочки и начало транскрипции его РНК. По данным современной литературы различают 6 способов проникновения вирусов в клетки: макропиноцитоз, три вида эндоцитоза (клатрин-зависимый, клатрин-независимый и холестерол-зависимый), с помощью образования кавеол (вогнутости плазматической мембраны) и подобный последнему механизм, зависимый от динамина. Как известно, взаимодействие вируса и клетки может происходить двумя путями: неспецифическим (инициируется при случайном столкновении, когда вирионы удерживаются на клеточной поверхности за счет электростатических сил) и специфическим (осуществляется при наличии на клеточной поверхности рецепторов, обеспечивающих прочное прикрепление вируса к клетке). После адгезии вирус проникает через плазмалемму и затем происходит перемещение его генома к определенным участкам цитоплазмы и ядра клетки, где инициируется процесс репродукции. 13. Этапы взаимодействия вируса с клеткой. На каком этапе реализуется генетическая информация вируса. 1.Адсорбция - пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина. Эффективность проникновения вируса связана большим количеством рецепторов. 2.Проникновение - путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза). 3.Освобождение нуклеиновых кислот - “раздевание” нуклеокапсида и активация нуклеиновой кислоты. 4.Реализация генетической информации. Т. е. синтез нуклеиновых кислот и вирусных белков - подчинение систем клетки хозяина и их работа на воспроизводство вируса. 5.Морфогенез вирусных частиц. Ассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком. 6.Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами. Ограничения, накладываемые клеткой на вирус. 1. В клетках про- и эукариот (кроме клеток растений) отсутствуют ферменты транскрипции-репликации вирусного РНК-генома. Вирус должен иметь свои ферменты в составе вириона или в закодированном виде в геноме. 2. В цитоплазме клеток нет ферментов для транскрипции ДНК вируса. Следовательно, клеточную ДНК-зависимую РНК-полимеразу могут использовать только ядерные ДНК-содержащие вирусы. 3. В клетках эукариот белоксинтезирующий аппарат приспособлен для трансляции моноцистронных РНК (не распознает внутренние участки инициации). Вирусы должны синтезировать или отдельные мРНК для каждого гена, или мРНК нескольких генов и кодирующий полипротеин, разрезаемый на отдельные белки. 4. В клетках прокариот возможна множественная внутренняя инициация трансляции на полицистронных матрицах. Для вирусов прокариот ограничение 3 снимается. 14. Супероболочка вирусов. Её происхождение, этапы формирования. Нуклеоид вирусов покрыт капсидом. Он состоит из капсомеров. А они в свою очередь из белковых субъединиц-протомеров. Капсомеры состоят в тесной связи с нуклеиновой кислотой, поэтому их вместе называют нуклеокапсидом. Капсид защищает НК, способствует переходу ее от одной клетки в другую. Вирусы, которые имеют только нуклеокапсид – простоорганизованные. У некоторых вирусов вирион имеет еще и дополнительную оболочку суперкапсид (пеплос). Такие вирусы- сложноорганизованные. По строению суперкапсид похож на цитоплазматическую мембрану клеток, что обусловлено его клеточным происхождением, так как он формируется в момент выхода вириона из пораженной клетки. Суперкапсид содержит в своем составе углеводы и липиды, а также небольшое количество белков, которые выполняют адресную функцию, то есть обеспечивают прикрепление вириона к поверхности клетки-хозяина. У большинства вирусов гликопротеиды формируют «шипы» на поверхности вирусной частицы. Шипы представляют собой морфологические субъединицы, построенные из нескольких молекул одного и того же белка. Вирусы гриппа имеют два типа шиповУ некоторых вирусов имеется также промежуточная оболочка – белковая мембрана. Особенности выхода из клетки оболочечных и безоболочечных вирусов. Различают два основных типа выхода вирусного потомства из клетки. Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки. Второй тип — почкование. Он присущ вирусам, имеющим суперкапсидную оболочку. На заключительном этапе сборки нуклеокапсиды сложно устроенных вирусов фиксируются на клеточной плазматической мембране, модифицированной вирусными белками, и постепенно выпячивают ее. В результате выпячивания образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки. Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции. 15. Морфогенез безоболочечных вирусов со спиральным типом симметрии (на примере ВТМ). 1. Образование структуры, состоящей из 2-х дисков, сформированных одинаковыми субъединицами. 2. Образование инициаторной петли РНК. 3. Встранивание петли РНК в отверстие диска (инициация сборки). 4. Элонгация (собственно сборка). Идет в 2-х направлениях вдоль цепи РНК, соповождается закручиванием спирали. 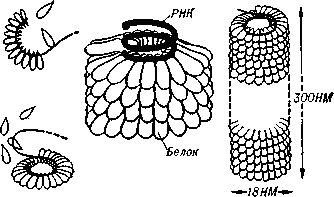 У оболочечных вирусов сначала формируются нуклеокапсиды, или сердцевины, которые затем покрываются белками наружных оболочек. Вирусы, имеющие оболочку (кроме вирусов оспы и реовирусов), формируются на клеточных мембранах. Все оболочечные вирусы млекопитающих со спиральным нуклеокапсидом, так же как некоторые вирусы с икосаэдрическим нуклеокапсидом (герпесвирусы, тогавирусы и ретровирусы), созревают при почковании через плазматическую мембрану, через внутреннюю цитоплазматическую мембрану или через мембрану ядра. 16. Морфогенез безоболочечных вирусов с изометрическим типом симметрии (на примере полиовируса). Все безоболочечные вирусы (Клеточные паразиты, центральный Капсид которых не окружен вирусными оболочками) позвоночных имеют кубическую симметрию. Структурные белки просто устроенных вирусов связываются спонтанно, образуя капсомеры, которые благодаря самосборке образуют капсиды, в которые упаковывается вирусная нуклеиновая кислота. Капсид полиовириона содержит 60 структурных единиц, состоящих из полипептидов VP1, 2, 3, 4. 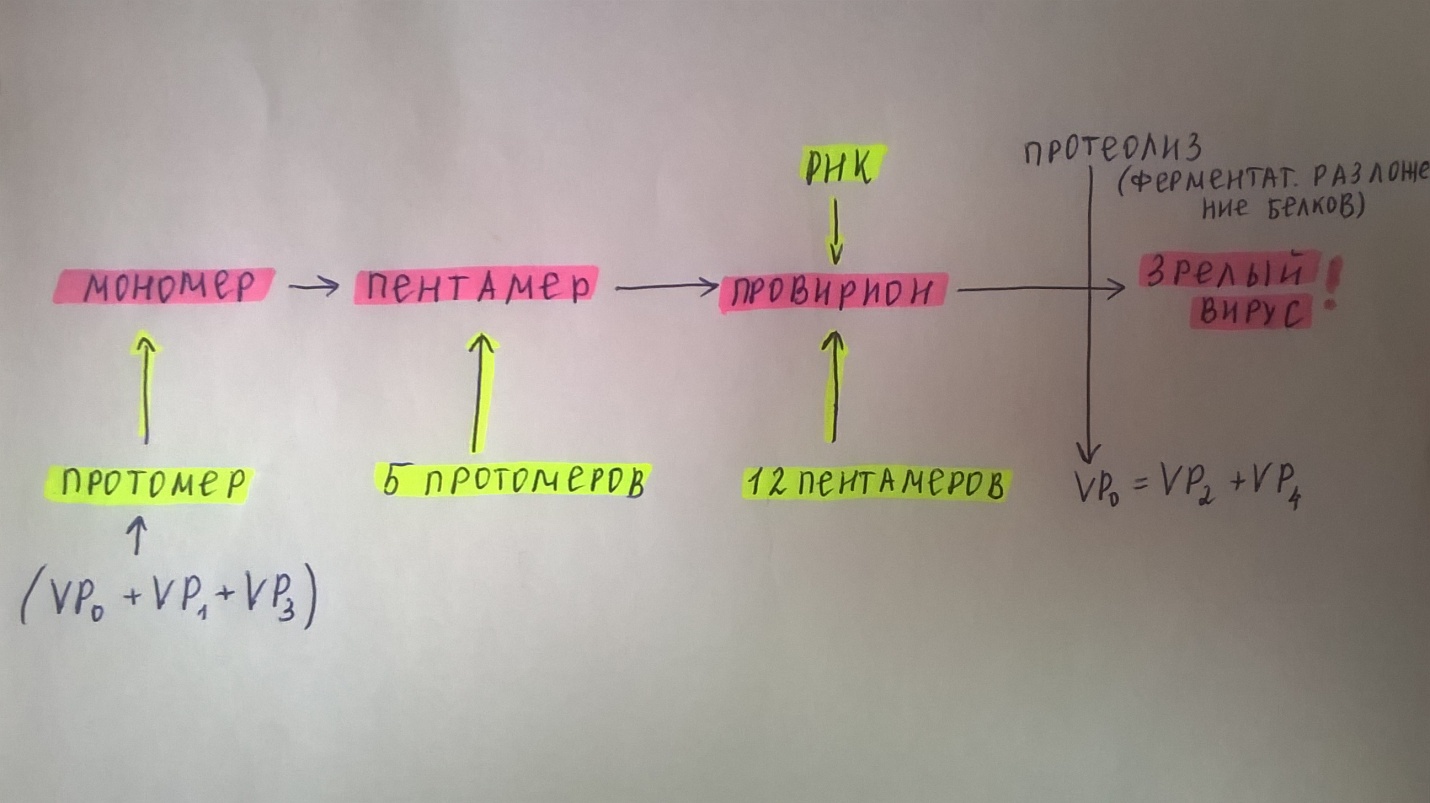 |