ЗАНЯТИЕ 4
Вопрос 1. Открытие ДНК и доказательства её генетической роли
ДНК была открыта в 1869 году Ф.Мишером.
1-е доказательство. В 1928 году Э.Гриффитсом было сделано предположение, что т.н. «вещество наследственности) находится в ядре. Гриффитс проделал следующий опыт: вначале он ввёл мышам непатогенные пневмококки – мыши выжили, затем биолог ввёл мышам патогенные микроорганизмы – мыши погибли. После этого Гриффитс ввёл мышам патогенные штаммы, убитые нагреванием – мыши выжили. Затем мышам были введены непатогенные микроорганизмы и убитые патогенные – мыши так же погибли. Гриффитс заключил, что живые микробы непатогенного штамма в присутствии клеток штамма патогенного приобретают наследственно закрепленные свойства патогенности (трансформируются). В последующем было доказано, что трансформация происходит не только в живом организме, но и in vitro, т.е. в пробирке.
2-е доказательство. В 1944 г. точно определили химическую природу «трансформирующего агента» и идентифицировали его как дезоксирибонуклеиновую кислоту. При обработке трансформирующего агента специфическими веществами, разрушающими ДНК, трансформация не осуществлялась. Таким образом, было получено прямое доказательство генетической роли ДНК.
3-е доказательство. Еще одним шагом в доказательстве генетической роли нуклеиновой кислоты является открытие правила эквивалентности, согласно которому в ДНК, выделенных из организмов различных видов, соотношение пуриновых и пиримидиновых оснований всегда одно и то же и составляет 1:1.
Вопрос 2. Строение нуклеотида ДНК
Нуклеотид ДНК состоит из
- азотистого основания (в ДНК 4 типа: аденин, тимин, цитозин, гуанин)
- моносахара дезоксирибозы
- фосфорной кислоты
Молекула нуклеотида состоит из трех частей — пятиуглеродного сахара, азотистого основания и фосфорной кислоты.
Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т. е. представляет собой пентозу. В зависимости от вида пентозы, присутствующей в нуклеотиде, различают два типа нуклеиновых кислот — рибонуклеиновые кислоты (РНК), которые содержат рибозу, и дезоксирибонуклеиновые кислоты (ДНК), содержащие дезоксирибозу. В дезоксирибозе — ОН-группа при 2-м атоме углерода заменена на атом Н, т. е. в ней на один атом кислорода меньше, чем в рибозе.
Вопрос 3. Типы нуклеотидов
В зависимости от пятиуглеродного сахара, содержащегося в нуклеотиде, выделяют РНК (рибоза) и ДНК (дезоксирибоза).
В состав нуклеотидов РНК могут входить азотистые основания: А, У, Г, Ц.
В состав нуклеотидов ДНК входят азотистые основания: А, Т, Г, Ц.
Вопрос 4. Первичная, вторичная и третичная структуры молекулы ДНК
Первичная структура – это последовательное расположение нуклеотидов в полинуклеотидной цепи ДНК. Между нуклеотидами имеется 3’,5’-фосфодиэфирная связь.
Вторичная структура ДНК представляет собой двойную спираль, состоящую из двух антипараллельных цепочек, закрученных вокруг одной оси. Цепочки соединяются водородными связями, которые образуются между азотистыми основаниями. Спираль ДНК закручивается вправо, общий виток 3,4нм, расстояние между цепочками 2нм.
Третичная структура – двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации с образованием суперспирали. Суперспиральная структура обеспечивает экономную упаковку огромной молекулы ДНК в хромосоме: вместо 8 см в вытянутой форме ДНК укладывается в 5 нм. Суперспирализация ДНК может быть нарушена разрывом в одной из цепей или в обеих цепях двойной спирали под действием ДНКазы.
Вопрос 5. Основные свойства ДНК
Универсальность - принципы построения ДНК для всех организмов одинаковы.
Специфичность определяется соотношением азотистых оснований, которое специфично для каждого вида.
Специфичность зависит от:
· количества нуклеотидов
· вида нуклеотидов
· расположения нуклеотидов в цепи ДНК.
Репликация. Репликация относится к реакциям матричного синтеза. Высокая надежность процесса репликации гарантирует практически безошибочную передачу генетической информации в ряду поколений.
Репарация - это свойство ДНК восстанавливать свою целостность, т.е. исправлять повреждения. Передача наследственной информации в неискаженном виде важнейшее условие выживания как отдельного организма, так и вида в целом. Большинство изменений вредны для клетки, они либо приводят к мутациям, либо блокируют репликацию ДНК, либо вызывают гибель клетки.
ДНК постоянно подвергается действию спонтанных (ошибки репликации, нарушение структуры нуклеотида и т.д.) и индуцированных (УФ – облучение, ионизирующая радиация, химические и биологические мутагены) факторов среды. В ходе эволюции выработалась система, позволяющая исправлять нарушения в ДНК – система репарации ДНК. В результате её активности на 1000 повреждений ДНК только одно приводит к мутациям. Повреждение - любое изменение ДНК, которое вызывает отклонение от обычной двуцепочечной структуры:
1) появление одноцепочечных разрывов;
2) удаление одного из оснований, в результате чего его гомолог остается неспаренным;
3) замещение одного основания в комплементарной паре другим, неправильно спаренным с основанием-партнером;
4) появление ковалентных связей между основаниями одной цепи ДНК или между основаниями на противоположных цепях.
Репарация может проходить до удвоения ДНК (дорепликативная репарация) и после удвоения ДНК (пострепликативная). В зависимости от характера мутагенов и степени повреждения ДНК в клетке идет световая (фотореактивация), темновая, SOS-репарация и др.
Считают, что фотореактивация идет в клетке, если повреждения ДНК вызваны естественными условиями (физиологические особенности организма, обычные факторы среды, в том числе - ультрафиолетовые лучи). Восстановление целостности ДНК при этом, происходит с участием видимого света: репаративный фермент активируется квантами видимого света, соединяется с поврежденной ДНК, разъединяет пиримидиновые димеры нарушенного участка и восстанавливает целостность нити ДНК.
Темновая репарация (эксцизионная)наблюдается после действия ионизирующей радиации, химических веществ и т.д. Она включает удаление поврежденного участка, восстановление нормальной структуры молекулы ДНК (рис.23). Для этого типа репарации необходима вторая комплементарная цепь ДНК. Темновая репарация многоступенчата, в ней участвует комплекс ферментов, а именно:
1) фермент, узнающий поврежденный участок цепи ДНК
2) ДНК – эндонуклеаза, делает разрыв в поврежденной цепи ДНК
3) экзонуклеаза удаляет измененную часть нити ДНК
4) ДНК – полимераза I синтезирует новый участок ДНК взамен удаленного
5)ДНК- лигаза сшивает конец старой нити ДНК с вновь синтезированной, т.е. замыкает два конца ДНК (рис.23). В темновой репарации у человека принимают участие 25 белков-ферментов.
При больших повреждениях ДНК, которые угрожают жизни клеток, включается SOS-репарация. SOS-репарация была открыта в 1974 году. Такой тип репарации отмечают после действия больших доз ионизирующей радиации. Характерная черта SOS-репарации - неточность восстановления первичной структуры ДНК, в связи с чем она получила название репарации, склонной к ошибкам. Главная цель SOS-репарации сохранить жизнеспособность клетки.
Нарушение в системе репарации могут приводить к преждевременному старению, развитию онкологических заболеваний, болезням аутоиммунной системы, гибели клетки или организма.
Вопрос 6. Механизм репликации
Репликация относится к реакциям матричного синтеза. Во время репликации каждая из двух цепей ДНК служит матрицей для образования комплементарной (дочерней) цепи. Протекает она в S-период интерфазы клеточного цикла. Высокая надежность процесса репликации гарантирует практически безошибочную передачу генетической информации в ряду поколений. Пусковым сигналом для начала синтеза ДНК в S-периоде являются специфические белки.
Репликон – это участок ДНК, где происходит репликация. Репликон содержит все необходимые, которые обеспечивают репликацию. Если процесс удвоения начался, он будет продолжаться до тех пор, пока весь репликон не будет удвоен.
У прокариот вся ДНК является одним репликоном.
У прокариот в процессе репликации принимает участие около 15 видов ферментов, а у эукариот – около 30-ти.
Ферменты репликации ДНК:
ДНК-полимеразы отвечают за рост дочерней цепи;
ДНК-праймаза необходима для действия ДНК-полимеразы;
ДНК – лигаза соединяет фрагменты Оказаки друг с другом за счет образования фосфодиэфирной связи;
ДНК – геликаза расплетает спираль ДНК, разрывает водородные связи между ними;
SSB – белки связываются с одноцепочечной ДНК и стабилизируют её.
Вопрос 7. Видовая специфичность молекулы ДНК
Число нуклеотндов и их последовательность в молекуле ДНК специфичны для каждого вида и частично — для каждой особи. Д. Уотсон ввел понятие о видовой специфичности ДНК. Коэффициентом видовой специфичности называют соотношение числа аденина и тимина к числу гуанина и цитозина.
В молекуле ДНК может быть записан практически любой объем наследственной информации, и у каждой особи эта запись уникальна и специфична. Генетический код значительно отличается даже внутри одного вида, лишь частично совпадая у родственников. И только однояйцевые близнецы имеют полностью идентичные хромосомы.
Иногда, вследствие мутаций (случайных изменений в структуре ДНК) организм получает свойства, не являющиеся нормой для его вида. Видовая специфичность молекул ДНК положена в основу современных методов диагностики инфекций – ПЦР (полимеразная цепная реакция). Проведение ПЦР-исследования позволяет по фрагменту ДНК, содержащемуся в крови или на слизистых оболочках человека, определить вид микроорганизма, вызвавший инфекционное заражение.
Вопрос 8. Уровни организации наследственного материала у эукариот
Наследственный материал эукариот больше по объему, чем у прокариот. Он расположен в основном в хромосомах, которые отделены от цитоплазмы ядерной оболочкой.
Генный уровень.
Элементарной функциональной единицей генетического аппарата является ген. Передачей генов в ряду поколений достигается материальная преемственность — наследование потомками признаков родителей.
Под признаком понимают отдельное качество или свойство, по которому организмы отличаются друг от друга.
Хромосомный уровень.
Гены клеток эукариот распределены по хромосомам, образуя ХРОМОСОМНЫЙ уровень организации наследственного материала. Этот уровень организации служит необходимым условием сцепления генов и перераспределения генов родителей у потомков при половом размножении (кроссинговер).
Геномный уровень.
Геном – вся совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Геном видоспецифичен, так как представляет собой необходимый набор генов, который обеспечивает формирование видовых характеристик организмов в ходе их онтогенеза.
Вопрос 9. Понятие о гене с молекулярной точки зрения. Свойства гена
Ген - это структурно-функциональная единица наследственности, представляющая собой участок молекулы ДНК. Гены бывают структурные и регуляторные. К структурным генам относятся гены транскрибирующие полипептидные цепи и РНК. К регуляторным относятся гены, участвующие в регуляции экспрессии генов.
Свойства генов:
- Мутация. Мутон – это минимальная единица мутирования. Может состоять из одной пары нуклеотидов.
- Рекомбинация. Рекон – единица рекомбинации. Минимальный размер рекона – две пары нуклеотидов.
- Дискретность – гены локализованы в определённых участках – локусах.
- Плейотропность – один ген отвечает за развитие нескольких признаков.
- Дозированность – несколько генов могут контролировать один признак. Чем больше генов, тем признак выражен ярче.
- Пенетрантность – свойство генов проявляться в фенотипе.
- Экспрессивность – степень фенотипического проявления гена.
- Специфичность – ген содержит информацию об определённом белке.
Вопрос 10. Структура гена прокариот
У прокариот различают:
1) структурные гены – они содержат информацию о последовательности аминокислот в молекуле белка;
2) функциональные гены – регулируют работу структурных генов;
3) структурные гены обычно расположены рядом и образуют один блок, который называется оперон. Оперон отвечает за строение молекулы белка и является структурной единицей наследственности прокариот;
4) в оперон входит промотор; промотор – это участок молекулы ДНК, к которому присоединяется РНК – полимераза. Кроме того, промотор определяет, с какой из двух цепей молекулы ДНК будет происходить транскрипция;
5) ген-оператор – регуляторный участок;
6) ген-терминатор – это ген, расположенный после структурного гена, на нём заканчивается процесс транскрипции;
7) ген-регулятор: он отвечает за кодировку структуры белка-репрессора.
Вопрос 11. Особенности структурной организации генов эукариот. Транскриптон
Ген (кодирующая часть) состоит из:
1) Экзонов.
2) Интронов.
Регуляторные участки гена содержат:
1)Стартовый кодон – место начала транскрипции.
2)Терминатор – место окончания транскрипции.
3) Лидерную последовательность.
4)Трейлерную последовательность.
5) Промотор.
6) Контролирующие зоны располагаются вблизи от обслуживаемого гена.
7) Модуляторы (энхансеры, сайленсеры) – располагаются вдали от гена.
Особенности кодирующей части гена:
1. Как правило, кодирующая область представлена не несколькими генами, а одним. Каждый ген у эукариот имеет свою регуляторную область.
2. Если в генах прокариот не кодирующие участки практически отсутствуют, то ген эукариот имеет мозаичное строение – в нём чередуются участки, несущие информацию о последовательности аминокислот в белке и не несущие её. Участки, несущие информацию носят название экзоны, не несущие называются интроны.
Особенности регуляторной части гена:
Регуляторные области – это участки ДНК на которых осаждаются белки-регуляторы. Их функция – регуляция транскрипции. Для простоты эти зоны подразделяют на два типа:
1) зоны, располагающиеся близко от гена, который они контролируют - контролирующие зоны.
2) зоны, располагающиеся далеко от контролируемого гена - модуляторы.
Участок ДНК, ограниченный промотором и терминатором, представляет собой единицу транскрипции - транскриптон (оперон у прокариот). В пределах каждого транскриптона копируется только одна из двух нитей ДНК, которая называется значащей или матричной.
Вопрос 12. Классификация генов
Все гены по функциям подразделяются на структурные и функциональные.
Структурные гены несут информацию о белках-ферментах и гистонах, о последовательности нуклеотидов в различных видах РНК.
Среди функциональных генов выделяют:
гены-модуляторы, усиливающие или ослабляющие действие структурных генов (супрессоры (ингибиторы), активаторы, модификаторы);
гены, регулирующие работу структурных генов (регуляторы и операторы).
Вопрос 13. Основные классы нуклеотидных последовательностей в ДНК клеток человека, их биологическая роль
В конце 60-х годов работами американских ученых Р. Бриттена, Э. Дэвидсона и других была открыта фундаментальная особенность молекулярной структуры генома эукариот – нуклеотидные последовательности разной степени повторяемости. Это открытие было сделано с помощью молекулярно-биологического метода изучения кинетики ренатурации денатурированной ДНК. Различают следующие фракции в геноме эукариот.
1. Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями. Как правило, это цистроны – структурные гены, кодирующие белки.
2. Низкочастотные повторы – последовательности, повторяющиеся десятки раз.
3. Промежуточные, или среднечастотные, повторы – последовательности, повторяющиеся сотни и тысячи раз. К ним относятся гены рРНК (у человека 200 на гаплоидный набор, у мыши – 100, у кошки – 1000, у рыб и цветковых растений – тысячи), тРНК, гены рибосомных белков и белков-гистонов.
4. Высокочастотные повторы, число которых достигает 10 миллионов (на геном). Это короткие ( 10 пн) некодирующие последовательности, которые входят в состав прицентромерного гетерохроматина.
Вопрос 14. Уровни упаковки наследственного материала при подготовке клеток к делению
-Нуклеосомный уровень- молекула ДНК взаимодействует с гистонами, формируя первичную нуклеосомную нить. Длина молекулы ДНК в результате упаковки сокращается в 6 раз;
- соленоид - второй уровень упаковкигенетического материала; образуется посредством конденсации нуклеосомной нити; уровень укладки достигает 40 раз;
- третий уровень конденсации – петли; на этом уровне генетический материала конденсируется в 600 - 1000 раз;
- четвертый уровень конденсации - метафазная хромосома- является результатом спирализации петель вокруг хромосомной оси; обеспечивается максимальный (10 000 раз) уровень упаковки генетического материала.
Вопрос 15. Расположение генов в хромосомах
Т. Морган установил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус.
Вопрос 16. Молекулярная организация хромосом
Каждая хромосома состоит из одной или двух молекул ДНК, которые, связываясь с гистоновыми и негистоновыми белками, образуют нуклепротеиновые комплексы с разным уровнем конденсации. В интерфазе хромосомы деконденсируются – они представлены в виде хроматина. В хроматине выделяют два типа участков: эухроматин, который транскрибируется и является генетически активным, и гетерохроматин, который не может быть транскрибирован и, таким образом, генетически неактивен. В метафазе клеточного деления хромосомы достигают максимальной конденсации, что позволяет точно идентифицировать хромосомы и изучить их морфологию.
Вопрос 17. Морфология метафазной хромосомы
Хромосомы лучше всего изучать во время метафазы митоза, т.к. в этой фазе они:
- располагаются в центре клетки, образуя метафазную пластинку;
- максимально конденсированы и легко различимы с использованием световой микроскопии;
- являются двухроматидными, а сестринские хроматиды соединены между собой в области центромеры, что позволяет различить их морфологию.
Морфологическими элементами метафазной хромосомы являются:
- 2 хроматиды;
- центромера;
- теломеры;
- вторичная перетяжка;
- спутник (сателлит);
- ломкие (фрагильные) участки.
Хроматида представлена одной линейной молекулой ДНК, связанной с гистоновыми и негистоновыми белками и максимально конденсированной. Метафазная хромосома состоит из двух сестринских хроматид.
Центромера, или первичная перетяжка, представляет собой участок хромосомы из ДНК и специальных центромерных белков. Положение центромеры в хромосоме постоянно и специфично для каждой хромосомы/пары гомологичных хромосом. Центромера делит хромосому на два плеча: проксимальное и дистальное. По положению центромеры хромосомы делятся на:
- метацентрические - центромера расположена посередине и плечи равные;
- субметацентрические - центромера несколько смещена к одному из концов, а плечи имеют разную длину;
- акроцентрические - центромера значительно смещена к концу хромосомы, из-за чего одно плечо намного короче другого.
Центромеры выполняют следующие функции:
- обеспечивают прикрепление хромосом к нитям веретена деления;
- обеспечивают разделение сестринских хроматид;
- точное и равное распределение генетического материала во время митоза.
Теломеры представлены специфическими последовательностями ДНК на концах хромосом в комплексе со специальными белками. Функции теломер:
- защищают концы хромосом от действия нуклеаз;
- предотвращают слипание концов хромосом;
- обеспечивают репликацию всей ДНК;
- контролируют процессы старения клеток;
- обеспечивают правильную конъюгацию гомологичных хромосом в мейозе.
Вторичные перетяжки - деспирализованные и слабо окрашенные участки повторяющейся ДНК. Вторичные перетяжки акроцентрических хромосом образуют область ядрышкового организатора.
Сателлиты – это терминальные участки коротких плеч акроцентрических хромосом, отделенные вторичной перетяжкой и состоящие из конститутивного гетерохроматина.
Ломкие (фрагильные) участки представляют собой деконденсированные сегменты хромосом, отличающиеся повышенной чувствительностью к действию мутагенных факторов, под влиянием которых в них легко происходят разрывы и, в результате этого, хромосомные перестройки.
Вопрос 18. Сателлитные и теломерные участки хромосом
Сателлиты - это терминальные участки коротких плеч акроцентрических хромосом отделенные вторичной перетяжкой и состоящие из конститутивного гетерохроматина; число и размеры сателлитов варьируют от индивида к индивиду.
Теломеры представлены специфическими последовательностями ДНК на концах хромосом в комплексе со специальными белками. Теломеры выполняют следующие функции:
- защищают концы хромосом от действия нуклеаз;
- предотвращают слипание концов хромосом;
- обеспечивают репликацию всей ДНК;
- предотвращают укорачивание хромосом благодаря активности теломеразы;
- контролируют процессы старения клеток и многоклеточного организма;
- регулируют фиксацию хроматина к ядерной мембране в интерфазе, обеспечивая тем самым нормальную архитектуру интерфазных хромосом;
- обеспечивают правильную конъюгацию гомологичных хромосом в мейозе.
Вопрос 19. Гетеро- и эухроматиновые участки хромосом
1. Гетерохроматин. Во время интерфазы определённые участки хромосом и целые хромосомы остаются компактными. Они образуют «глыбки» интенсивно окрашенные и, как правило, прилежащие к мембране ядра. Гетерохроматин неактивен в отношении транскрипции. Существует две формы гетерохроматина: факультативный и конститутивный
факультативный гетерохроматинбывает гетерохроматичным только временами. Он информативен, содержит гены с которых считывается наследственная информация, когда гетерохроматин переходит в эухроматическое состояние. Образуется при спирализации одной из двух гомологичных хромосом. Типичным примером служит тельце полового хроматина, образуемого одной из двух Х-хромосом соматических клеток женских особей человека и млекопитающих. Функциональная роль факультативного гетерохроматина заключается в компенсации снижении дозы определенного гена (например, появление промежуточного признака при явлении неполного доминирования у гетерозигот Аа, влияет на экспрессивность проявления наследственных признаков в фенотип), определяет тканеспецифичность.
конститутивный гетерохроматин– отличается высокоспирализованным состоянием, которое сохраняется на протяжении всего мит. цикла. Он занимает постоянные участки в гомологичных хромосомах – это фрагменты околоцентромерных, теломерных участков хромосом, Не содержит структурных генов (нетранскрибируемый); Его роль не ясна, но по видимому он выполняет опорную функцию.
2. Эухроматин (разрыхленный) - имеет менее компактную организацию, деспирализуется в конце митоза, образует слабоокрашенные нитчатые структуры содержит структурные транскрибируемые гены.
В КАЖДОЙ ХРОМОСОМЕ СВОЙ ПОРЯДОК РАСПОЛОЖЕНИЯ ЭУ- И ГЕТЕРОХРОМАТИНА, ЧТО ИСПОЛЬЗУЕТСЯ ДЛЯ ИДЕНТИФИКАЦИИ ОТДЕЛЬНЫХ ХРОМОСОМ В ЦИТОГЕНЕТИКЕ.
Вопрос 20. Соотношение понятий хроматин и хромосома
Хроматин и хромосомы представляют собой разновидности генетических комплексов, способных переходить друг в друга. Их химическая организация сходна между собой.
Хромосома. Хромосомой принято называть элемент, входящий в состав ядра клетки. Хромосома участвует в формировании структуры ядра. Это хранилище ДНК, а значит и информации наследственного типа об организме в целом. Линейный порядок расположения генов характеризует хромосомы. Хромосому формируют хроматиды. Хроматиды представлены при этом парой продольных субЪединиц. Каждая из входящих в обозначенную пару хроматид по своему строению и структуре абсолютно аналогична другой хроматиде. Основу хроматиды составляет молекула ДНК, представленная в единичном экземпляре. Теломеры выступают конечными участками хроматид.
Хроматин. Хроматин является веществом, входящим в состав хромосомы. Его можно выделить из ядер клеток растений или животных. Для хроматина характерна способность интенсивно окрашиваться ядерными красителями. Когда клетка начинает делиться, хроматин претерпевает процесс формирования в различимые структуры определённого типа, находящиеся в составе хромосом.
Общность обозначенных понятий заключается в следующем:
Процесс спирализации хроматина, в ходе которого и происходит конечное образование хромосомы.
Кроме того, и хроматин, и хромосома являются двумя структурно-функциональными состояниями, которые несут в себе наследственный материал.
Отличия. Хромосомы, как элементы ядра клетки, имеют следующие отличительные особенности, расходящиеся с особенностями хроматина:
Способность к самовоспроизведению.
Наличие индивидуальной составляющей, связанной со структурными и функциональными свойствами.
Возможность сохранять индивидуальные характеристики в ряду нескольких поколений.
Хроматин от хромосомы можно отличить по следующим параметрам:
Является комплексом, переставленным сочетанием РНК, ДНК и белков.
Представляет собой вещество хромосом.
Располагается внутри клеток эукариот.
Является составным фрагментом нуклеотида у прокариот.
С помощью своего состава наделён способностью реализовывать генетическую информацию.
Реализует репликацию и репарацию ДНК.
Хромосомы и хроматин можно различать ещё и по внешнему облику. Хроматин представлен глыбками, гранулированными элементами и нитчатыми структурами, в целом можно сказать, что это уплотнённые участки хромосом.
Хромосомы, будучи единицей генетического материала, наделены плотной структурой.
Различна реакция хромосомы и хроматина на окрашивание. Хроматин поддаётся воздействию только некоторых красителей, к ним можно отнести кармин и гематоксилин. Хромосомы окрашиваются интенсивно, чутко реагируя на красящий элемент.
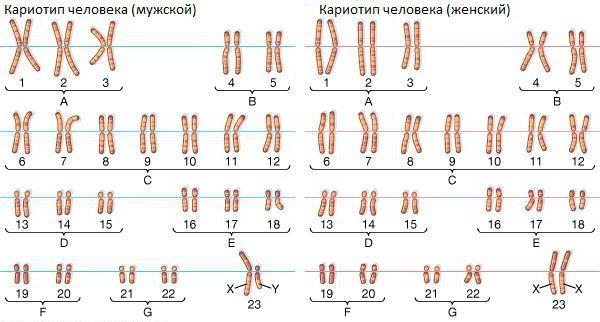
Вопрос 21. Кариотип, его видовая специфичность
Кариотип — совокупность признаков (число, размеры, форма и т.д.) полного набора хромосом, присущий клеткам опр биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Видовая специфичность кариотипа заключается в том, что каждый вид имеет определенные число, форму и размеры хромосом. Каждая хромосома внутри одной группы сцепления занимает определенный, свой локус. Если говорить о парах гомологичных хромосом с их гомологичными локусами, то они отличаются лишь характером аллельных генов, но не по форме и размерам. Хромосомы имеют определенный набор генов, и каждый ген занимает определенное место относительно других (локус-это место внутри хромосомы, в котором находится ген). Это также очень специфичный признак не только для каждой хромосомы из генофонда вида, но, тем более, для хромосом из набора различных видов.
Вопрос 22. Характеристика кариотипа человека
У человека нормой является наличие хромосом, количество которых равно 46. Из них 22 пары являются аутосомами и две — это половые хромосомы.
У представительниц женского пола они обозначаются как ХХ, у представителей мужского пола — ХУ. Главная особенность хромосомного набора — это видовая специфичность кариотипа. Функции хромосом заключаются в том, что каждая из них является носительницей генов, которые отвечают на наследственность.
Нормальный мужской кариотип — это кариотип 46, ХУ. Нормальный женский кариотип выглядит как кариотип 46, ХХ.
В характеристике кариотипа вначале указывается общее число хромосом и набор половых хромосом. Затем (при наличии мутаций) указываются геномные, а после – хромосомные мутации. Большое диагностическое значение для идентификации хромосом имеют хромосомные маркеры (chromosome landmarks) – полосы, наиболее четко выделяющиеся при дифференциальной окраске и разделяющие хромосому на районы. На некоторых полосах удается локализовать определенные гены генетической карты хромосомы.
Вопрос 23. Сущность Денверской классификации хромосом человека и её основные принципы
В 1960 году в городе Денвере, США, состоялась конференция по хромосомам человека. Решением конференции стала так называемая Денверская классификация хромосом человека. Данная система руководствуется принципами:
1.Все аутосомы человека пронумерованы по порядку от 1 до 22 по мере уменьшения их длины, половым хроматидам присвоены обозначения X и Y.
2.Хромосомы кариотипа разбиты на 7 групп с учетом положения центромер, наличия спутников и вторичных перетяжек на хроматидах.
3. Для упрощения классификации используется центромерный индекс (ЦИ), который рассчитывается делением длины короткого плеча на всю длину хромосомы и выражается в процентах.
Денверская классификация хромосом включает семь групп, в которых аутосомы расположены по порядку нумерации, но распределены неравномерно по количеству.
Группа A. Пары 1—3. Длинные хромосомы, хорошо отличимые друг от друга. В 1 и 3 парах положение перетяжки метацентрическое, во 2 паре – субметацентрическое.
Группа В. Пары 4 и 5. Хромосомы короче предыдущей группы, первичная перетяжка расположена субметацентрически (близко к середине).
Группа С. Пары 6—12, X-хромосома. Хромосомы среднего размера, все неравноплечие субметацентрические, сложно идивиндуализируемые. Идентична по размеру и форме аутосомам группы, репликацию заканчивает позже других.
Группа D. Пары 13—15. Хромосомы в группе средних размеров с практически краевым положением первичной перетяжки (акроцентрические), имеют спутники.
Группа E. Пары 16—18. Короткие хромосомы, в 16 паре равноплечие метацентрические, в 17 и 18 - субметацентрические.
Группа F. Пары 19 и 20. Короткие метацентрики, практически неотличимы друг от друга.
Группа G. Пары 21 и 22, Y-хромосома. Короткие хромосомы со спутниками, акроцентрические. Имеют незначительные отличия строения и размеров. Немного длиннее других хромосом группы, на длинном плече имеется вторичная перетяжка.
Вопрос 24. Особенности организации генома эукариот
Геном - совокупность генов гаплоидного набора хромосом. Ген- это линейный участок молекул ДНК, в котором закодирована последовательность а/к одной полипептидной цепи молекул белка. Большинство генов эукариот имеют мозаичное строение, т. е. состоят и чередующихся кодирующих (экзоны) и не кодирующих (интроны) участков.
Последовательности:
Уникальные, т.е. последовательности, представленные в одном экземпляре или немногими копиями.
Среднеповторяющиеся – последовательности, повторяющиеся сотни и тысячи раз.
Высокоповторяющиеся, число которых достигает 10 миллионов на геном.
Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль над временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме.
Полученные к настоящему времени данные свидетельствуют, что экспрессия генов эукариот регулируется на уровне транскрипции, процессинга РНК в ядре и стабильности мРНК. Кроме того, было показано, что на экспрессию эукариотических генов оказывают влияние амплификация и перестройка генов.
Вопрос 25. Проект «Геном человека»
В 1986 году группа ученых в США начала работу над проектом, позднее названным «Геном человека». Цель этого проекта заключалась в том, чтобы представить в виде карты полную последовательность ДНК человека.
Так, в 1995 году Институтом изучения генома в Роквилле, штат Мэриленд, была опубликована первая полная последовательность ДНК живого организма — бактерии Haemophilus influenzae.
В 1996 году был определен первый геном эукариотической клетки. Каждое такое достижение требовало определения все более и более длинной последовательности и было важной вехой на пути к определению генома человека.
В июне 2000 года Крейг Вентер и Фрэнсис Коллинз, руководители проекта «Геном человека», осуществлявшегося в Национальных институтах здоровья США, объявили о событии, названном ими «первой сборкой генома человека». По существу, это была первая реконструкция полного генома человека.
В 2003 г. этот проект был успешно завершен. Ученые описали все 25 000 генов, присутствующих в хромосомах каждой клетки. За это время были созданы базы ДНК из образцов генов десятков тысяч людей.
Френсис Коллинз, руководитель программы «Геном человека» в США, директор Национального института исследований генома человека, дает такой прогноз результатов геномных исследований:
2020 г.
На рынке появляются лекарства от сахарного диабета, гипертонии и других заболеваний, разработанные на основе геномной информации.
Терапия онкологических заболеваний, прицельно направленная на свойства раковых клеток.
Фармакогеномика становится общепринятым подходом для создания многих лекарств.
Изменение способа диагностики психических заболеваний, появление новых способов их лечения, изменение отношения общества к таким заболеваниям.
Демонстрация безопасности генной терапии на уровне зародышевых клеток при помощи технологии гомологичной рекомбинации.
2030 г.
Определение последовательности нуклеотидов всего генома отдельного индивида станет обычной процедурой.
Каталогизированы гены, участвующие в процессе старения.
Проводятся клинические испытания по увеличению максимальной продолжительности жизни человека.
Лабораторные эксперименты на человеческих клетках заменены экспериментами на компьютерных моделях.
2040 г.
Все общепринятые меры здравоохранения основаны на геномике.
Определяется предрасположенность к большинству заболеваний (при или до рождения).
Доступна эффективная профилактическая медицина с учетом особенностей индивида.
Болезни определяются на ранних стадиях путем молекулярного мониторинга.
Для большинства заболеваний доступна генная терапия.
Замена лекарств продуктами генов, вырабатываемых организмом при ответе на терапию.
Средняя продолжительность жизни достигнет 90 лет.
Серьезные дебаты о возможности для человека контролировать свою собственную эволюцию.
Вопрос 26. Эволюция генома
Общие принципы организации наследственного материала у про- и эукариот свидетельствуют в пользу единства их происхождения от общего предка. Однако эволюция названных типов клеток шла по разным направлениям после их дивергенции от общего предка.
Первоначальный отбор шел на способность к самосохранению через самовоспроизведение. В соответствии с рассмотренным предположением позднее некоторые участки ДНК приобрели функцию кодирования, т.е. стали структурными генами, совокупность которых на определенном этапе эволюции составила первичный генотип. Проявление возникших кодирующих последовательностей ДНК привело к формированию первичного фенотипа, который подвергался естественному отбору.
Важным моментом в рассматриваемой гипотезе является предположение о том, что существенным компонентом первых клеточных геномов была избыточная ДНК, способная реплицироваться, но не несущая функциональной нагрузки в отношении формирования фенотипа. Предполагают, что разные направления эволюции геномов про- и эукариот связаны с различной судьбой этой избыточной ДНК. Избыточность ДНК создавала возможность расширения объема кодирующих нуклеотидных последовательностей за счет некодирующих, обеспечивая возникновение многих вариантов решения проблемы формирования жизнеспособного фенотипа.
|
 Скачать 177.5 Kb.
Скачать 177.5 Kb.