Физа шизойда. 7. Современное представление о центре кровообращения. Роль различных отделов центральнойнервной системы в регуляции кровообращения
 Скачать 7.46 Mb. Скачать 7.46 Mb.
|

|
7. Современное представление о центре кровообращения. Роль различных отделов центральнойнервной системы в регуляции кровообращения   Сердечно-сосудистые центры ствола головного мозга следует: Сосудосуживающий центр (также называемый С1) расположены в верхнем мозговом веществе и нижнем мосту. Эфферентные нейроны этого сосудодвигательного центра входят в состав симпатической нервной системы и синапсов в спинной мозг, затем симпатические ганглии и наконец, на органы-мишени, вызывая сужение сосудов в артериолах и венулах. Кардиоускорительный центр. Эфферентные нейроны из кардиоускорительного центра также являются частью симпатическая нервная система и синапсы в спинной мозг, симпатические ганглии и, наконец, сердце. В сердце последствия этой деятельности повышенная скорострельность узла SA (для увеличения частота сердечных сокращений), увеличение скорости проводимости через атриовентрикулярный узел и повышенная сократимость. Центр сердечного замедления. Эфферентные волокна из центр сердечных замедлителей являются частью пара- симпатическая нервная система: они путешествуют в блуждающий нерв и синапс на узле SA, чтобы уменьшить частота сердцебиения. Сердечно-сосудистые центры ствола головного мозга расположены в ретикулярной формации продолговатого мозга и нижних одна треть моста. Эти центры функционируют в координированная мода, получение информации о крови давление от барорецепторов, а затем направление изменение активности симпатической и парасимпатической нервной системы, чтобы скорректировать кровяное давление, как нужный. Как описано выше, артериальное давление определяется барорецепторами. Также в каротидном синусе и дуге аорты. Афферентная затем информация об артериальном давлении отправляется в продолговатый мозг через языкоглоточный (CN IX) и блуждающий (CN X) нервы. Эта информация интегрирована в ядра солитарного тракта, которое затем направляет изменения в деятельности ряда сердечно-сосудистых центров. Эти сердечно-сосудистые центры тонически активны, а ядро одиночного тракта просто направляет через центры увеличивается или уменьшается отток симпатических и парасимпатической нервной системы. Парасимпатический отток является следствием блуждающий нерв на узле SA, чтобы уменьшить частоту сердечных сокращений. Симпатический отток состоит из четырех компонентов: влияние на узел SA для увеличения частоты сердечных сокращений, эффект на сердечную мышцу для повышения сократительной способности и инсульта объем, воздействие на артериолы, вызывающее вазокон-стрикцию и увеличение TPR, а также воздействие на вены вызывают сужение вен и уменьшают ненапряженный объем. 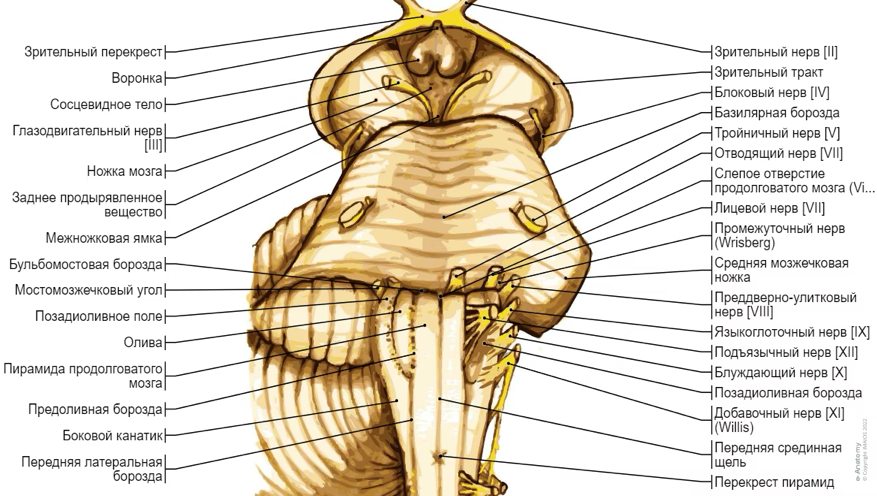 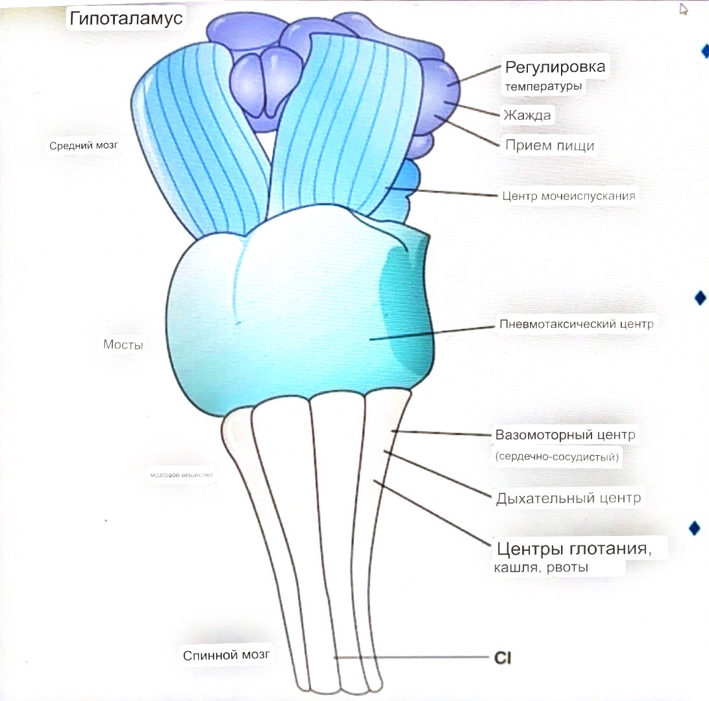 Все черепные нервы, кроме одного, отходят от вентральная поверхность мозга; блоковый (IV) нерв берет начало от дорсальной поверхности среднего мозга.  (  Лучше не получалось) 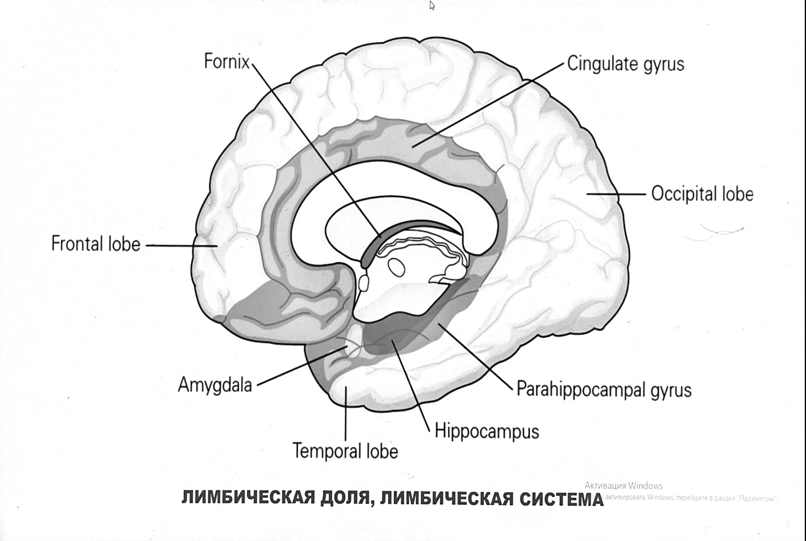 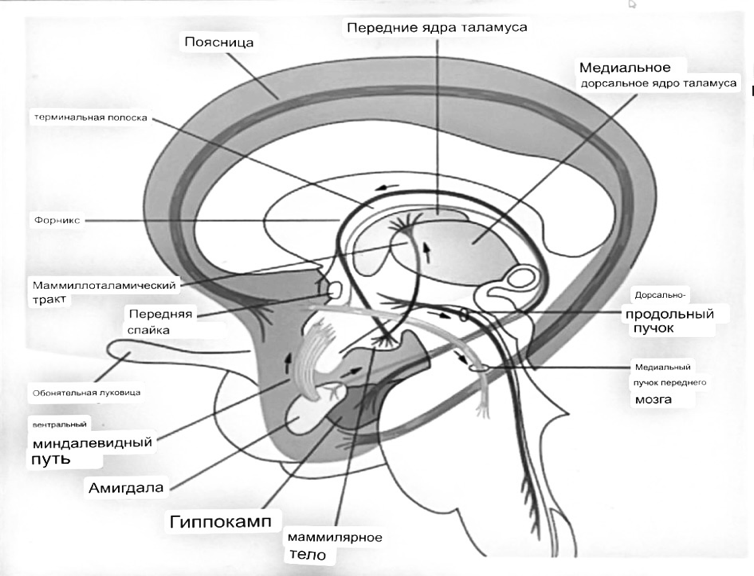 Выше-примитивная корковая ткань Н 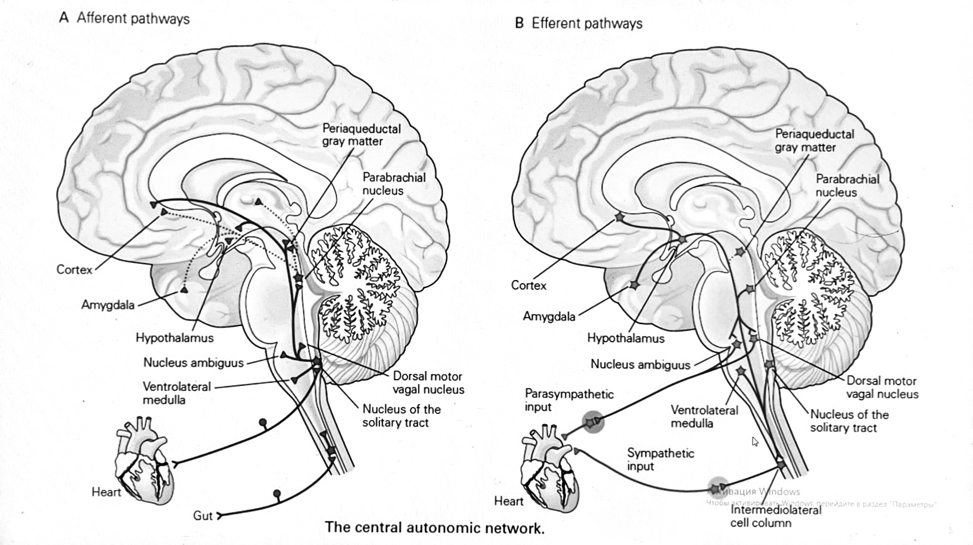 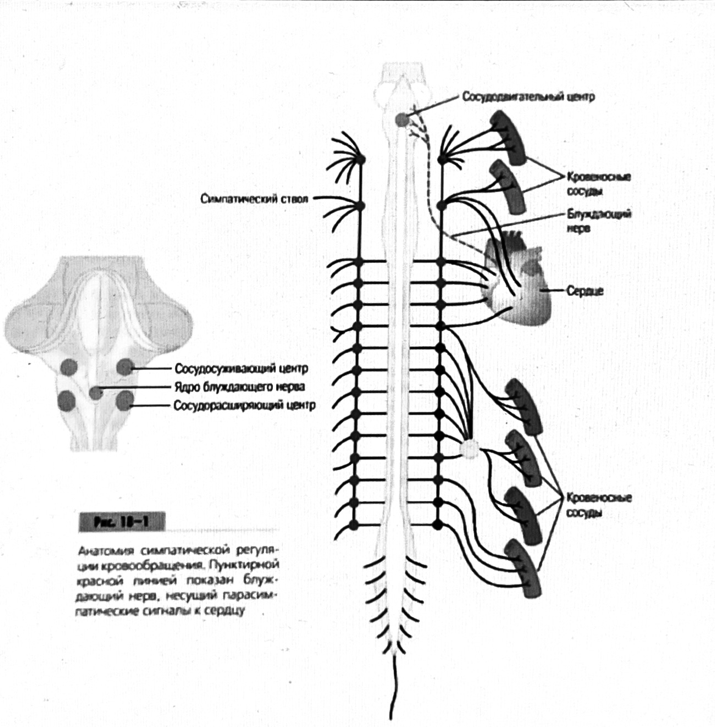 иже-г 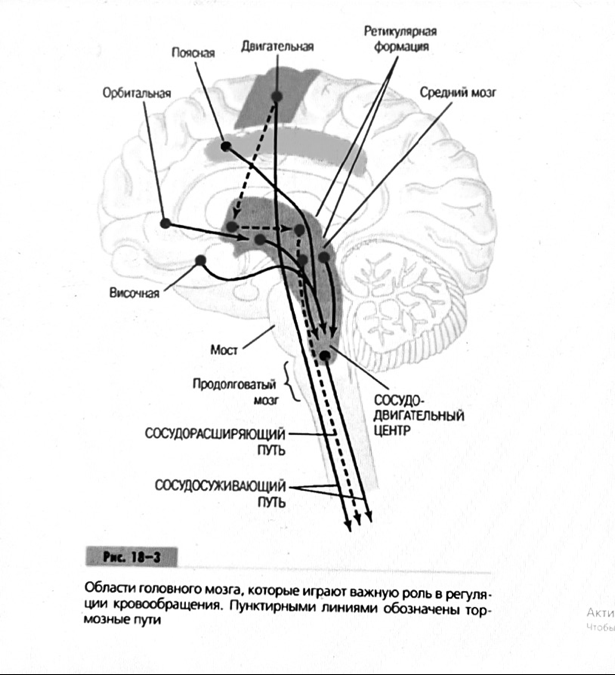 иппокамп,миндалевидное тело  Вегетативная нервная система и артериальные барорецепторные рефлексы синоатриальные и парасимпатические нервы иннервируют синоатриальный (СА) и атриовентрикулярный (АВ) узлы. Мио- кардиальный отдел иннервируется симпатическими нервами. Артериальные и венозные сосуды на большей части тела иннервируется симпатическими нервами, тогда как парасимпатическая нервная система иннервирует сосуды в половых органов и желудочно-кишечного тракта. Вегетативная эфферентная активность регулируется барорецепторным рефлексом. в ответ на изменение артериального давления, выявляемое барорецепторами каротидного синуса и дуги аорты. проиллюстрирована реакция на изменение позы. АХ, ацетилхолин; АВ, атриовентрикулярный; ЦНС, центральная нервная система; Е, адреналин; MAP, среднее артериальное давление; НЭ, норадреналин; СА, синоатриальный 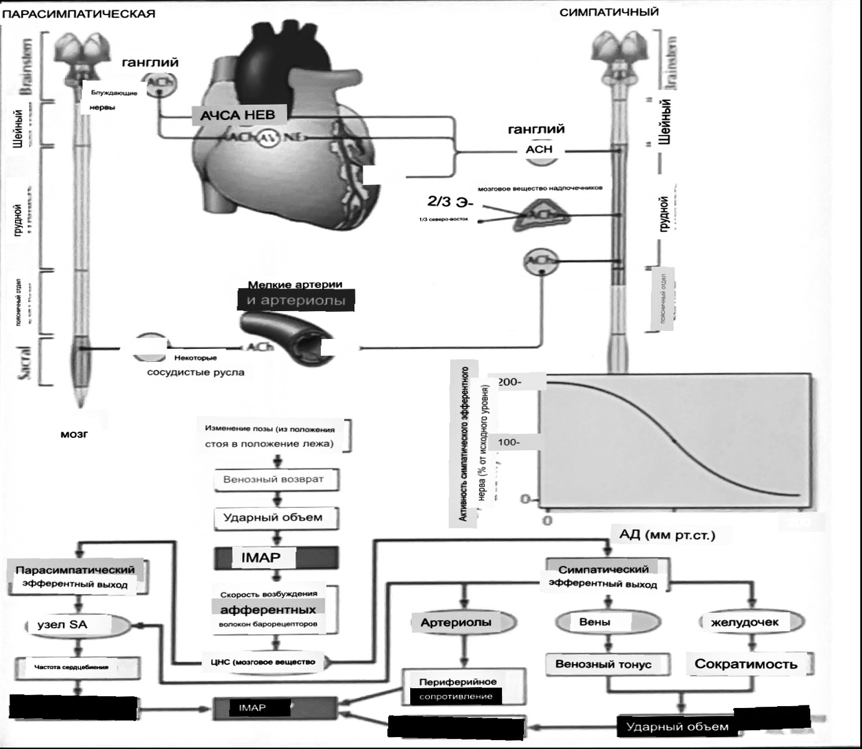 Общая схема центральной регуляции.На рис.7.27 представлена общая схема организации центральных звеньев нервной регуляции кровообращения. Сердечно-сосудистый центр продолговатого мозга состоит из двух зон (А и Б), одна из которых определяет сужение сосудов (А'), ускорение и усиление сердцебиений (А"), тогда как другая — расширение сосудов (БI) и замедление частоты сокращений сердца (БII). Тонически активные нейроны зоны А непосредственно или через промежуточные нейроны возбуждает прегангли-онарные и ганглионарные симпатические нейроны, иннервирующие сердце и сосуды, и в то же время тормозят нейроны зоны Б. Последние активируются сигналами барорецепторов сердечно-сосудистой системы. Через короткие внутрицентральные пути нейроны 331 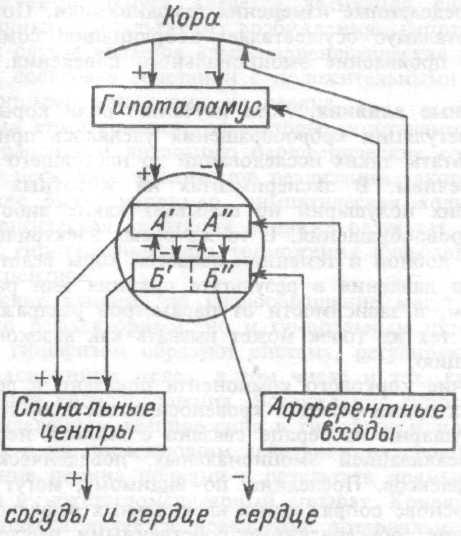 Рис.7.27. Схема организации центральных звеньев нервной регуляции сердечно-сосудистой системы. Объяснение в тексте. зоны Б тормозят нейроны зоны А, а посредством нисходящих тормозных путей — и преганглионарные симпатические нейроны. Кроме того, через моторное ядро блуждающих нервов нейроны зоны Б влияют на ганглионарные парасимпатические нейроны сердца. В результате обе зоны (А и Б), получая основную сигнализацию от рецепторов сердечно-сосудистой системы, являются основой поддержания кровообращения, сочетательно подвергая сердце тоническому контролю тормозных вагусных и возбуждающих симпатических волокон, тогда как постоянный контроль сосудов обеспечивается только симпатическими вазоконстрикторными волокнами. Афферентная нервная система является источником сигналов собственных и сопряженных рефлексов. Вход первых ограничен продолговатым мозгом, причем барорецепторы сердечно-сосудистой системы связаны с зоной Б, а хеморецепторы — с зоной А. С последней связаны также и некоторые афференты сопряженных рефлексов (от рецепторов кожи, мышц и внутренних органов), хотя организация этих связей известна лишь в общих чертах. Поскольку сердечно- сосудистый центр продолговатого мозга регулирует основные параметры системной гемодинамики, даже будучи отключенным от связей с вышележащими нервными структурами, он функционирует как истинный "центр", обладающий значительной независимостью. Следовательно, супрабульбарные структуры признаются несущественными ни для прямой тонической регуляции кровообращения, ни для осуществления сердечно-сосудистых рефлексов. В то же время при общих реакциях организма (поведенческих, эмоциональных, стрессорных и др.) кора мозга, гипоталамус и другие супрабульбарные структуры могут вмешиваться в деятельность сердечно-сосудистой системы, изменяя путем нисходящих (модулирующих) влияний возбудимость бульбарного центра. Вазомоторные центры спинного мозга самостоятельного значения в регуляции кровообращения не имеют. Действующими элементами спинального уровня являются лишь симпатические преганглионар-ные нейроны, которые входят в состав эфферентного звена буль-барных рефлекторных дуг. Только в случае травм, приводящих к утрате связей продолговатого мозга со спинным, последний, спустя некоторое время, приобретает способность осуществлять вазомоторные рефлексы на импульсы спинномозговых афферентов. 8. Поддержание постоянства артериального давления в регуляции системного кровообращения.Срочные (рефлекторные) механизмы регуляции артериального давления. Рефлексы с основных рефлексогенных зон. Значение баро-, хемо-и волюмо-рецепторов. Собственные и сопряжённые рефлексы. Условно-рефлекторная регуляция кровообращения.  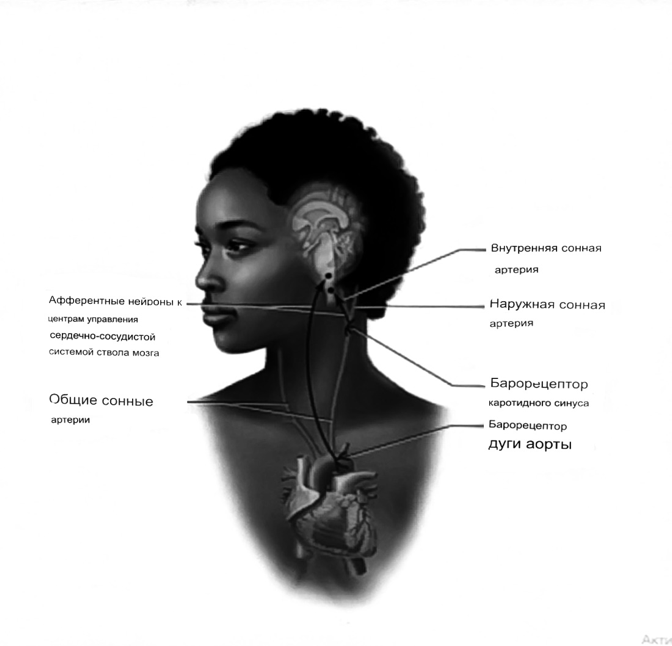 Значение баро-, хемо-и волюмо-рецепторов.   Возбуждение хеморецепторов аортальной и синокаротидной рефлексогенных зон возникает при уменьшении напряжения О2, увеличении напряжения СО2 и концентрации водородных ионов, т.е, при гипоксии, гиперкапнии и ацидозе. Импульсы от хеморецепторов поступают по тем же нервам, что и от барорецепторов, в продолговатый мозг, но непосредственно к нейронам прессорного отдела симпатического центра, возбуждение которого вызывает сужение сосудов, усиление и ускорение сердечных сокращений и как следствие — повышение АД. В результате кровь быстрее поступает к легким, углекислый газ обменивается на кислород. Изменения деятельности сердечно-сосудистой системы способствуют устранению отклонений от нормы газового состава крови. Однако эффект невелик, так как увеличение АД осуществляется главным образом за счет сужения сосудов и лишь частично — в результате стимуляции деятельности сердца. Такой же эффект наблюдается и при нарушении кровоснабжения мозга любого происхождения (недостаточности деятельности сердца, нарушении мозгового кровообращения). Если степень нарушения кровоснабжения мозга выражена сильно, АД можети повыситься до 200 мм рт.ст. и больше. 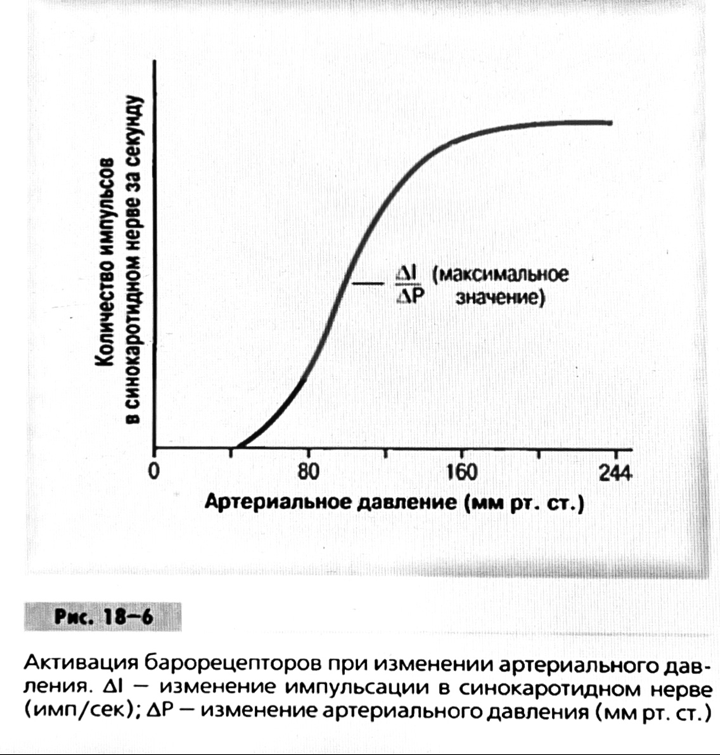 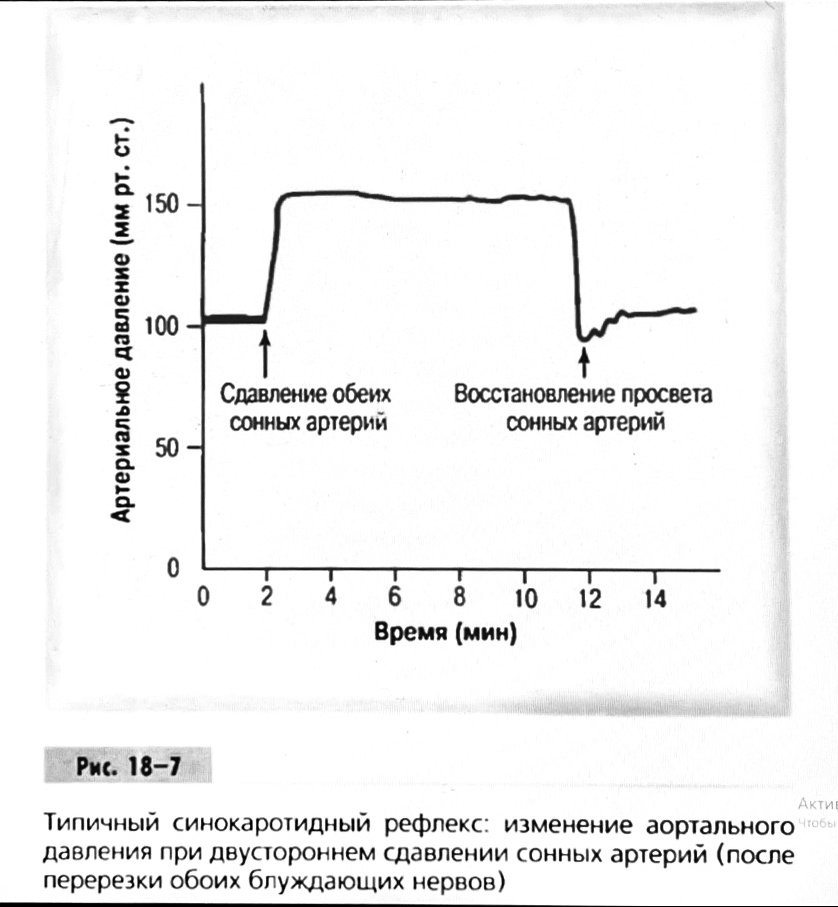  Барорецептор. частота стрельбы меняется с изменением артериальное давление. (a) Влияние изменения среднего артериальное давление (MAP) в начале действия потенциалы афферентными нейронами каротидной синус. Этот эксперимент проводится путем накачивания кровь непульсирующим образом через изолированный каротидный синус, чтобы можно было установить давление внутри него при любом желаемом значении. (b) Активация потенциала действия барорецептора частота колеблется в зависимости от давления. Увеличивать при пульсовом давлении увеличивается общее действие потенциальная частота даже при нормальном АД. 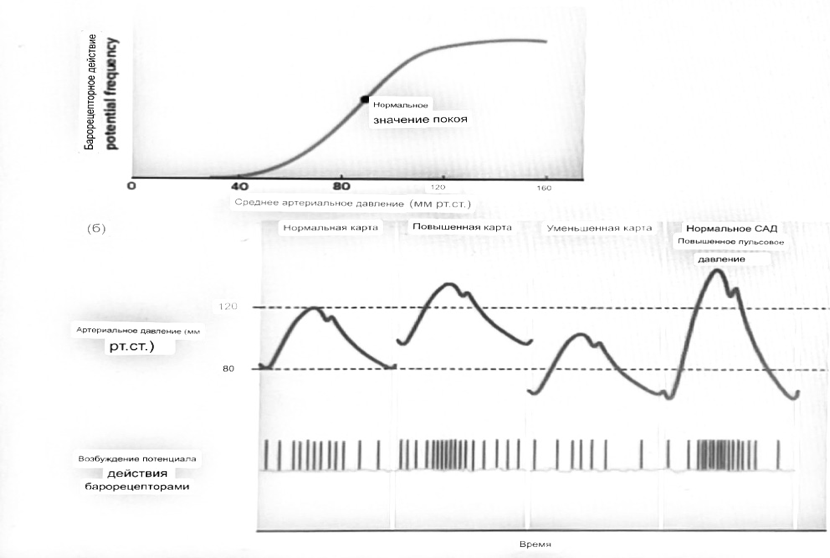 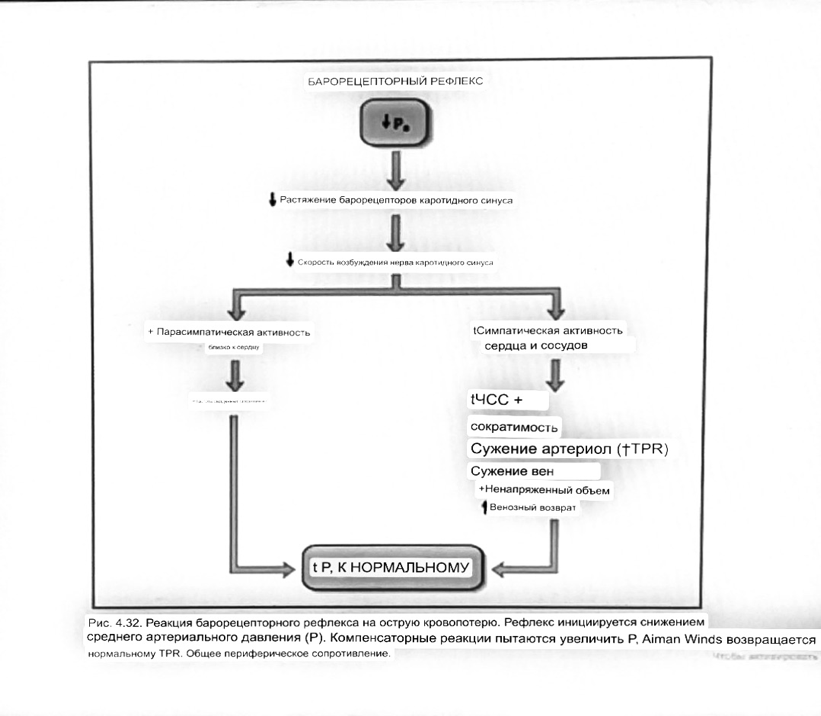 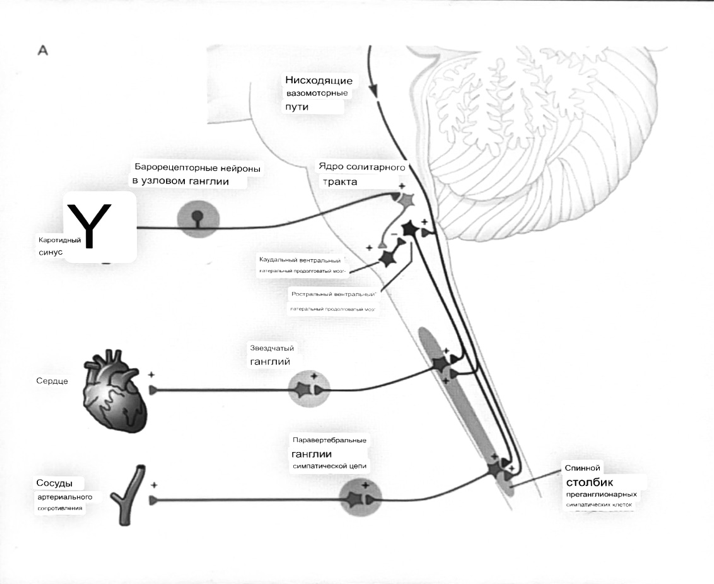  Барорецепторный рефлекс ведет себя как отрицательный обратная связь с усилением. A. Артериальное кровяное давление воспринимается барорецепторами, типом рецепторный нейрон растяжения в каротидном синусе у основания мозг. После интеграции в медулию эта информация про- обеспечивает контроль сердечно-сосудистой системы с отрицательной обратной связью. Симпатический компонент схемы включает в себя выходы которые стимулируют насосную способность сердца (сердечный выброс) за счет увеличения частоты сердечных сокращений и силы сокращений. В кроме того, симпатическая стимуляция вызывает сокращение артерий. что повышает гидравлическое сопротивление току крови. Вместе последствия увеличения сердечного выброса и увеличения сосудистого резистентность к повышению среднего артериального давления. Важно, тормозные проекции от каудального отдела к ростральному вентральному латеральный продолговатый мозг создает отрицательную обратную связь, так что увеличение артериальное давление угнетает симпатическую активность, в то время как снижение повышает симпатическую активность. Хотя опущено для простота, парасимпатические нейроны в кардиальном узле также способствуют рефлексу, создавая тормозной сердечный вход который функционально антагонистичен симпатическому пути (см. Рисунок 49-98). Во время барорецепторных рефлексов парасимпа- Поэтому активность в сердце увеличивается за счет гипер- напряжение и снижается при гипотонии, 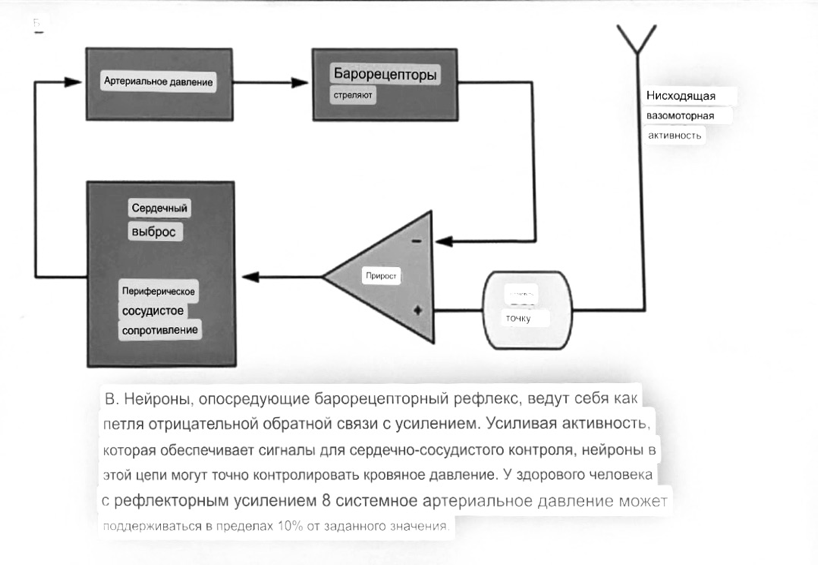 Срочные (рефлекторные) механизмы регуляции артериального давления. Механизмы быстрого реагирования- это рефлекторная регуляция АД с помощью изменений работы сердца и тонуса (просве1 та) сосудов. Эти реакции срабатывают в течение нескольких секунд. Причем, в случае повышения АД работа сердца тормозится, тонус сосудов уменьшается - они расширяются. И то, и другое ведет к снижению (нормализации) АД. Если же давление снижается, то деятельность сердца увеличивается, а сосуды сужаются, что ведет к увеличению - нормализации АД. Включаются в реакцию и емкостные сосуды. В случае повышения АД тонус емкостных сосудов уменьшается, что ведет к задержке крови в венах, уменьшению притока крови к сердцу и уменьшению выброса крови сердцем. В случае снижения АД тонус емкостных сосудов возрастает, что ведет к увеличению возврата крови к сердцу и возрастанию выброса сердцем крови. Рецепторы, воспринимающие изменения кровяного давления, барорецепторы (точнее, рецепторы растяжения) рассеяны по всему кровеносному руслу, но имеются их скопления: в дуге аорты и в области каротидного синуса (главные сосудистые рефлексогенные зоны), в сердце (предсердиях, желудочках, коронарных сосудах), легком, в стенках крупных грудных и шейных артерий. В перечисленных участках имеются многочисленные барорецепторы,а в дуге аорты и каротидном синусе - баро- и хеморецепторы.Хотя принцип работы рефлексогенных зон одинаков, их значение в регуляции АД несколько различается. 1.Основные рефлексогенные зоны сосудистого русла: дуга аорты (барорецепторы); каротидный синус (место разветвления общей сонной артерии на наружную и внутреннюю) (хеморецепторы); устье полых вен (механорецепторы); емкостные кровеносные сосуды (волюморецепторы). 2.Внесосудистые рефлексогенные зоны. Основные рецепторы рефлексогенных зон сердечнососудистой системы: Барорецепторы и волюморецепторы, реагирующие на изменение АД и объема крови (относятся к группе медленно адаптирующихся рецепторов, реагируют на деформацию стенки сосуда, вызванную изменением АД и/или объема крови). Барорефлексы. Повышение АД приводит к рефлекторному урежению сердечной деятельности, снижению ударного объема (парасимпатическое влияние). Падение давления вызывает рефлекторное увеличение ЧСС и повышение УО (симпатическое влияние). Рефлексы с волюморецепторов. Уменьшение ОЦК ведет к увеличению ЧСС (симпатическое влияние). 1.Хеморецепторы, реагирующие на изменение концентрации кислорода и углекислого газа крови. При гипоксии и гиперкапнии ЧСС увеличивается (симпатическое влияние). Избыток кислорода вызывает уменьшение ЧСС. 2.Рефлекс Бейнбриджа. Растяжение устий полых вен кровью вызывает рефлекторное увеличение ЧСС (торможение парасимпатического влияния). Рефлексы с внесосудистых рефлексогенных зон. Классические рефлекторные влияния на сердце. 1.Рефлекс Гольца. Раздражение механорецепторов брюшины вызывает урежение сердечной деятельности. Такой же эффект при механическом воздействии на солнечное сплетение, сильном раздражении Холодовых рецепторов кожи, сильных болевых воздействиях (парасимпатическое влияние). 2.Рефлекс Данини-Ашнера. Надавливание на глазные яблоки вызывает урежение сердечной деятельности (парасимпатическое влияние). 3. Двигательная активность, несильные болевые раздражения, активация тепловых рецепторов вызывают увеличение ЧСС (симпатическое влияние). Начало формы ВОЛЮМОРЕЦЕПТОРЫ: ПОЛЫЕ ВЕНЫ, ПРЕДСЕРДИЯ, УШКИ СЕРДЦА РЕФЛЕКС БЕЙНБРИДЖА: РАСТЯЖЕНИЕ ПОЛЫХ ВЕН И ПРАВОГО ПРЕДСЕРДИЯ УВЕЛИЧИВАЮТ ЧСС (ПОВЫШЕНИЕ СИМПАТИЧЕСКОГО ТОНА) (НО! СЛИШКОМ БОЛЬШОЙ ОБЪЕМ ПРИТЕКАЮЩЕЙ КРОВИ СНИЖАЕТ ЧСС) РЕФЛЕКС ПАРИНА: РАЗДРАЖЕНИЕ РЕЦЕПТОРОВ ЛЕГОЧНОЙ АРТЕРИИ ИНГИБИРУЮТ РАБОТУ СЕРДЦА И РАСШИРЯЮТ СОСУДЫ БОЛЬШОГО КРУГА КРОВООБРАЩЕНИЯ (ПРЕДУПРЕЖДЕНИЕ ОТЕКА ЛЕГКИХ) РЕФЛЕКС КИТАЕВА: РАЗДРАЖЕНИЕ РЕЦЕПТОРОВ ЛЕВОГО ПРЕДСЕРДИЯ СУЖАЕТ СОСУДЫ МАЛОГО КРУГА КРОВООБРАЩЕНИЯ (ПРЕДУПРЕЖДЕНИЕ ОТЕКА ЛЕГКИХ) РЕФЛЕКС ГЕНРИ-ГАУЭРА: РАЗДРАЖЕНИЕ РЕЦЕПТОРОВ ПРЕДСЕРДИЙ И УШКОВ ПРИВОДИТ К СНИЖЕНИЮ СЕКРЕЦИИ АДГ ГИПОТАЛАМИЧЕСКИМИ ЯДРАМИ. В ПОЧКАХ УМЕНЬШАЕТСЯ РЕАБСОРБЦИЯ ВОДЫ, ДИУРЕЗ УВЕЛИЧИВАЕТСЯ, ОБЪЕМ КРОВИ УМЕНЬШАЕТСЯ (НЕЙРО-ЭНДОКРИННЫЙ РЕФЛЕКС) Собственные и сопряженные рефлексы  СОБСТВЕННЫЕ (СИСТЕМНЫЕ) РЕФЛЕКСЫ КРОВООБРАЩЕНИЯ — рефлексы, определяющие регуляцию взаимоотношений в пределах деятельности одной физиологической системы — сердечно-сосудистой (В. Н. Черниговский). При относительно небольших интенсивностях раздражения рецепторов они возникают без сопровождения другими рефлексами. Напр., изменение давления в каротидных синусах (см.) прежде всего сказывается на кровообращении (собственный рефлекс кровообращения) и уже потом (при увеличении интенсивности раздражения) на дыхании (сопряженный рефлекс). Переход от собственных рефлексов к сопряженным часто совершается постепенно и последние в ряде случаев могут рассматриваться как определенная фаза в развитии ряда последовательных цепей рефлексов, вызываемых раздражением данной интероцептивной зоны. Пороги раздражения рецепторов для собственных рефлексов во всех случаях ниже порогов для сопряженных рефлексов. Рефлекторные изменения сердечной деятельности могут проявляться при активации любой сенсорной системы. Исходя из этого, академик В. Н. Черниговский в 1960 г. предложил выделять три категории кардиальных рефлексов: 1) собственные рефлексы, вызываемые раздражением рецепторов сердечно-сосудистой системы; 2) сопряженные рефлексы, обусловленные активностью любых других рефлексогенных зон; 3) неспецифические эффекты раздражения, которые воспроизводятся в условиях физиологического эксперимента, а также при патологии. Однако далеко не каждая воспроизводимая в условиях эксперимента нейрогенная реакция сердца имеет значение для кровообращения в интактном организме, а многие висцеральные рефлексы оказывают на сердце побочное или неспецифическое действие. Наибольшее значение в регуляции кровообращения имеют собственные рефлексы сердечно-сосудистой системы, которые в физиологической литературе классифицируются по трем группам: 1) васкуло-кардиальные, 2) кардиокардиальные и 3) кардиоваскулярные рефлексы. Васкуло-кардиальные рефлексы.( эффект Бейнбриджа)Наибольшее значение среди них имеют барорефлексы, возникающие в ответ на изменения системного артериального давления. . Павлов, Используя в качестве воздействий стимуляцию и холодовое отключение сердечных нервов, апноэ, введение атропина и усиленное потребление жидкости, И. П. Павлов убедился, что эти воздействия вызывают существенные изменения деятельности сердца, но при этом непропорционально малые изменения артериального давления. Как уже отмечалось, адекватность кровоснабжения организма требует соблюдения по крайней мере трех необходимых условий. Первое условие – достаточная величина минутного объема кровообращения (3–6 л/мин с возможностью увеличения при нагрузках в 5–6 раз). Это условие обеспечивается миогенными, нейрогенными и гуморальными механизмами регуляции деятельности сердца. Вторым условием является распределение минутного объема крови в соответствии с текущими потребностями организма. Данное условие обеспечивается местными миогенными и гуморально-метаболическими механизмами. Третьим условием является обеспечение оптимального градиента артериального давления. От этого показателя зависит линейная скорость тока эритроцитов в сосудах. Составляя в аорте 150 см/c, в капиллярном русле различных органов она снижается до 0,2–2,7 мм/c.( В результате каждый эритроцит проводит в капилляре от 250 до 750 мс, что представляет собой отрезок времени, сравнимый с тем, который необходим для газообмена с окружающей тканью. Линейная скорость эритроцитов при функциональных нагрузках может быть увеличена без ущерба для газовой функции крови примерно в два раза. При дальнейшем увеличении линейной скорости крови возможно уменьшение артериовенозной разницы по кислороду, отражающее ухудшение качества кровоснабжения тканей.) Таким образом, оптимальное снабжение клеток и тканей кислородом возможно в ограниченном диапазоне линейных скоростей крови в микроциркуляторном русле. Уровень системного артериального давления зависит от интенсивности сердечной деятельности (сердечный компонент) и гидравлического сопротивления сосудистого русла (сосудистый компонент). Коссвенная оценка первого показателя производится по результатам сопоставления величин венозного притока, ударного (систолического) объема и частоты сердечных сокращений, а второго – по исчисленному параметру – общему периферическому сопротивлению сосудов (ОПСС). Как говорилось выше, ОПСС рассчитывается по упрощенной формуле Пуазейля как частное от деления среднего системного артериального давления на сердечный выброс. Несмотря на ограничения в приложении закона Пуазейля к потоку крови (турбулентность кровотока, неньютоновские жидкостные свойства крови, переменный диаметр сосудистого русла, непостоянная вязкость крови), данный параметр удобен для непрерывной регистрации и позволяет судить о величине нагрузки на сердце, поскольку имеет ту же размерность, что и гидравлическое сопротивление. Однако он в принципе не может адекватно отражать сосудистый компонент регуляции артериального давления, поскольку последнее в не меньшей степени зависит от величины сердечного выброса, нежели от просвета сосудов. В этом заключается методологическая ошибка многих исследований системной гемодинамики, в которых ОПСС рассматривается как самостоятельный физиологический показатель. Рефлекторный механизм регуляции артериального давления реализуется при участии системных артериальных барои хеморецепторов, роль которых была изучена благодаря исследованиям В. Бейлиса (1893), Х. Геринга (1923–1927) и К. Хейманса (1930-е гг.). У высших млекопитающих и человека основные барорецепторные зоны расположены в области дуги аорты, плечеголовной и сонной артерий . Ведущую роль в обеспечении рефлекторного контроля кровообращения играют барорецепторы дуги аорты и синокаротидных зон (области разделения общих сонных артерий на внутренние и наружные). Подобные барорецепторы представляют собой медленно адаптирующиеся механорецепторы растяжения, реагирующие на деформацию сосудистой стенки при изменениях кровяного давления. Пороговое значение абсолютной величины внутрисосудистого (систолического) давления, необходимого для получения устойчивой импульсации по волокнам типа А, составляет около 60–120 мм рт. ст., а в волокнах типа С – 120–150 мм рт. ст. Уровень насыщения, достигаемый при повышении давления, для обеих групп барорецепторов не превышает 200 мм рт. ст., после чего они перестают реагировать на дальнейшее повышение давления. Абсолютный порог чувствительности рецепторов дуги аорты составляет 90–120 мм рт. ст., то есть выше, чем у рецепторов каротидного синуса, где он равен примерно 60–80 мм рт. ст. Сосудистые барорецепторы реагируют на изменение не только средней величины артериального давления, но и частоты, а также амплитуды его пульсовых колебаний. Каждая пульсовая волна вызывает импульсное растяжение сосудистой стенки, которое возбуждает рецепторы. При нормальном среднем артериальном давлении импульсная активность в синусовом нерве носит фазный характер, усиливаясь при каждой систоле и уменьшаясь или исчезая после дикротического подъема на пульсовой кривой. При повышении артериального давления частота разрядов каждого волокна увеличивается, а при понижении уменьшается. Импульсная активность в аортальных нервах аналогична таковой в синусных нервах. Вспышки импульсной активности в аортальных нервах возникают несколько раньше, чем в синусных, что определяется скоростью распространения пульсовой волны по магистральным артериям. Афферентные пути от барорецепторов сердечно-сосудистой системы заканчиваются в продолговатом мозге в области ядра одиночного пути (nucleus tractus solitarius ). Таким образом, подавляющее большинство рефлекторных изменений артериального давления осуществляется с обязательным вовлечением ствола головного мозга. Совокупность структур ретикулярной формации каудальной части ствола головного мозга, при участии которых происходит замыкание рефлексов сердечно-сосудистой системы, традиционно обозначается как бульбарный (медуллярный) кардиоваскулярный центр (рис. 44).. Уже говорилось, что этот центр содержит нейроны в составе полисенсорных и полимодальных ядер ретикулярной формации (ядро одиночного пути, парамедианное ядро шва, дорзальное ядро вагуса) и одиночные нейроны, диффузно расположенные на вентральной поверхности продолговатого мозга. Сердечный компонент нейрогенной регуляции обеспечивается тормозными (депрессорными) влияниями, которые реализуются парасимпатической системой через преганглионарные нейроны и их аксоны в составе блуждающего нерва, а также нисходящими возбуждающими влияниями на спинальные преганглионарные симпатические нейроны, участвующие в иннервации сердца. Сосудистый компонент формирования артериального давления реализуется через внутриорганный нервный аппарат артериальных сосудов, в составе которого преобладают аксоны и терминали адренергических симпатических нейронов. ( 1 подпункт) Другой компонент симпатоадреналового механизма регуляции артериального давления обеспечивается рефлекторными влияниями на секрецию адреналина и норадреналина клетками мозгового вещества надпочечников в плазму крови. Эта секреция находится под контролем преганглионарных симпатических волокон, благодаря чему в плазме крови в любой момент времени имеется физиологически значимая концентрация катехоламинов. В случае повышения величины кровяного давления в аорте или в каротидном синусе рефлекторно усиливаются тонические тормозные влияния блуждающих нервов на сердце. При этом отмечено и некоторое торможение тонической активности симпатических сердечных нервов, но по сравнению с усилением парасимпатических тормозных влияний на сердце его роль менее значима. В результате уменьшаются частота сердечных сокращений, сократимость миокарда и сердечный выброс, что приводит к снижению артериального давления. Уменьшение давления в аорте и каротидном синусе вызывает, напротив, уменьшение импульсной активности в аортальном и синусовом нервах, что приводит к рефлекторной активации бульбарных центров регуляции системы кровообращения и усилению тонических симпатических влияний на сердце, а также сосуды. В результате возрастают частота сердечных сокращений, сократимость миокарда, сердечный выброс и артериальное давление. Указанные сдвиги деятельности сердца могут проявляться в ответ на пережатие сонных артерий у животных в эксперименте (синокаротидный прессорный рефлекс) (рис. 45). Барорецепторные зоны, участвующие в регуляции сердечной деятельности, имеются и в других отделах сосудистой системы. Например, в легочных и почечных артериях обнаружены рецепторы, функции и электрофизиологические характеристики которых близки к барорецепторам, расположенным в дуге аорты. Поэтому в случае повышения давления в легочной артерии также рефлекторно замедляется ритм сердца и снижается сердечный выброс, что способствует уменьшению давления в легочной артерии. В 1938 г. К. Хеймансом в сосудистой системе обнаружены хеморецепторные зоны, которые находятся в каротидных тельцах, расположенных дистальнее бифуркации сонных артерий, и в аортальном тельце, расположенном в области дуги аорты. Обе рефлексогенные зоны реагируют на изменение напряжения кислорода в артериальной крови и участвуют таким образом в регуляции дыхания. Возбуждение рефлексогенных зон при снижении парциального давления кислорода в артериальной крови вызывает повышение частоты сердечных сокращений, а также констрикцию периферических сосудов. Этот механизм отличается исключительно высокой чувствительностью. Так, например, у человека увеличение частоты сердечных сокращений отмечено уже при снижении напряжения кислорода в артериальной крови всего на 3 %. Хеморецепторы каротидных и аортальных телец являются особенно значимыми в рефлекторной регуляции деятельности сердца в условиях острой гипоксической гипоксии (высокогорье, задержка дыхания при нырянии). (+к 1 подпункту) Параллельно с симпатоадреналовой системой в регуляции артериального давления принимает участие еще один механизм – ренин-ангиотензиновая система. Запуск этой системы осуществляется посредством секреции почками протеолитического фермента ренина. Синтез ренина увеличивается посредством местного рефлекса при снижении артериального давления. Под действием ренина от α-глобулина плазмы ангиотензиногена отщепляется биологически инертный декапептид ангиотензин I. В капиллярах легких под действием ангиотензин-превращающего фермента (АПФ) от ангиотензина I отщепляются еще две аминокислоты. В результате образуется октапептид ангиотензин II. Последний обладает двойным физиологическим действием: он является сильным вазоконстриктором и стимулирует секрецию корой надпочечников альдостерона, тем самым уменьшая диурез и увеличивая объем циркулирующей крови. Последующее спонтанное отщепление от молекулы ангиотензина II еще одной аминокислоты приводит к образованию ангиотензина III, обладающего сосудосуживающим действием. В отличие от катехоловых аминов, ангиотензин II и ангиотензин III действуют прежде всего на артериальные сосуды, лишь незначительно влияя на сердечную деятельность и почти не изменяя тонус вен. Кардиокардиальные рефлексы. Как было отмечено в главе 1, в стенках сердечных камер и в эпикарде расположено большое количество различных по модальности рецепторов. Однако наибольшее значение для рефлекторной регуляции деятельности сердца имеют механорецепторы, расположенные в предсердиях в месте впадения в них полых и легочных вен, а также в левом желудочке. Такие рецепторы в физиологической литературе называются также волюморецепторами. Исследованиями установлено, что А-рецепторы реагируют на изменение напряжения стенок предсердий, а В-рецепторы возбуждаются при их пассивном растяжении при увеличении венозного притока крови к сердцу. Как правило, в ответ на растяжение предсердий рефлекторно усиливаются тормозные влияния блуждающих нервов на сердце, что приводит к снижению частоты сердечных сокращений, сократимости миокарда и сердечного выброса. Данный рефлекс представляет наглядный пример отрицательной обратной связи в нейрогенной регуляции деятельности сердца. Так, увеличение объема циркулирующей крови, приводя к повышению венозного возврата, вызывает повышение давления в правом предсердии. В результате растяжения предсердий рефлекторно уменьшается частота сердечных сокращений, сократимость миокарда, что приводит к снижению сердечного выброса и венозного притока. Р. Бейнбриджем был обнаружен рефлекс, при котором в ответ на увеличение давления в правом предсердии при исходно низкой частоте сокращений сердца у собак происходило увеличение частоты сердечных сокращений. В физиологических исследованиях 1960–1970-х гг. в экспериментах на разных видах теплокровных животных показано, что направленность сдвигов частоты сердечных сокращений в ответ на растяжение предсердий зависит от исходной частоты ритма сердца, который косвенно зависит от тонической активности блуждающих нервов. При низкой исходной частоте сердечных сокращений (60–80 уд/мин) в ответ на увеличение давления в правом предсердии развивается рефлекторная тахикардия, тогда как при высокой исходной частоте ритма сердца (120–180 уд/мин) при увеличении давления в правом предсердии имеет место замедление частоты сердечных сокращений, то есть брадикардия. В клинической кардиологии считается, что при сердечной недостаточности, когда на фоне снижения сократимости миокарда давление в правом предсердии повышается, рефлекс Бейнбриджа, вызывая тахикардию, способствует увеличению сердечного выброса. Однако в условиях тахикардии сердце работает неэффективно, так как продолжительность диастолы уменьшается, что ухудшает кровенаполнение сердца и коронарный кровоток. В результате сократимость миокарда еще более снижается и недостаточность насосной функции сердца прогрессирует. Из физиологической литературы известно также, что при возбуждении волюморецепторов предсердий происходит увеличение диуреза и уменьшение объема циркулирующей крови. Рефлекс называется по имени авторов, впервые открывших его, – кардиоренальным рефлексом Генри – Гауэра. В исследованиях 1970–1980-х гг. показано, что увеличение диуреза в ответ на растяжение предсердий является следствием усиления выработки кардиомиоцитами предсердий натрийуретического пептида (см. гл. 7). Кардиоваскулярные рефлексы. Возник в связи с проблемой механизмов регуляции венозного притока крови к сердцу. Э. Нил высказал гипотезу о рефлекторной регуляции тонуса венозных сосудов с участием рецепторов, расположенных в правом предсердии. Исследования, выполненные позднее в школе академика Б. И. Ткаченко, выявили разнонаправленность реакций венозных сосудов, возникающих в ответ на раздражение рецепторов предсердий. Эти данные свидетельствуют о неспецифичности направленности кардиоваскулярных рефлексов. В 1983 г. английский физиолог С. Роте высказал предположение о наличии сопряженного влияния симпатических нервов на сердце и венозные сосуды, обеспечивающего при «мобилизации» резервного объема крови из вен увеличение сократимости миокарда. Однако остаются невыясненными афферентные пути этих рефлексов. Вероятно, в этом случае невозможно выделить единственный афферентный путь. Сопряженныекардиальные рефлексы представляют собой реакции сердца в ответ на раздражение рефлексогенных зон, расположенных вне системы кровообращения. К значимым в клиническом отношении сопряженным рефлексам относится ряд реакций, названных по именам описавших их авторов: – рефлекс Гольца; – рефлекс Тома – Ру; – рефлекс Данини – Ашнера. Рефлекс Гольцапроявляется в форме брадикардии или даже полной остановки сердца в ответ на раздражение механорецепторов брюшины. Возможность возникновения этого рефлекса учитывают при проведении оперативных вмешательств в брюшной полости, когда во избежание рефлекторной остановки сердца в область брыжейки вводят новокаин или применяют блокатор М-холинорецепторов – атропин, который устраняет парасимпатические влияния на сердце. Рефлекс Тома – Ру– брадикардия вплоть до остановки сердца при сильном давлении или ударе в эпигастральную область может явиться в боксе и других единоборствах причиной преходящей недостаточности мозгового кровообращения с потерей сознания. Кроме того, рефлекторная остановка сердца может наблюдаться при резком охлаждении кожи области живота. Именно такую природу нередко имеют несчастные случаи при нырянии в холодную воду. Рефлекс Данини – Ашнерапроявляется в форме брадикардии при надавливании на глазные яблоки. Он является примером сопряженного соматовисцерального кардиального рефлекса. Данный рефлекс раньше использовался в неотложной медицине в комплексе мер для купирования приступов пароксизмальной тахикардии. Ротовая полость также является важной рефлексогенной зоной. При охлаждении ротовой полости в ответ на раздражение чувствительных окончаний тройничного нерва у человека может быть рефлекторное снижение частоты сокращений сердца и расширение коронарных артерий. Например, препарат валидол, содержащий масло мяты, помещают под язык при приступе стенокардии. В результате раздражения рецепторов ротовой полости приступ стенокардии купируется. К числу сопряженных кардиальных рефлексов относятся также все условные рефлексы, влияющие на деятельность сердца. Примером является так называемая предстартовая лихорадка – состояние, которое наблюдается у спортсменов, скаковых лошадей и борзых собак перед началом соревнований. Оно представляет собой следствие активации симпатоадреналовой системы, которая возникает в ответ на комплекс условных раздражителей, сопутствующих проведению спортивных мероприятий. Характерным примером условнорефлекторных изменений частоты сердечных сокращений у человека является «синдром белого халата», когда при посещении врача возрастает частота сердечных сокращений сердца и повышается артериальное давление. Формирование такого условного рефлекса происходит еще при первом посещении врача ребенком. Если, например, врач выполняет инъекцию препарата, которая сопровождается болью, стрессорной реакцией, приводящей к тахикардии, то возникает условный рефлекс: врач – условный раздражитель, болевой стимул – безусловный. В дальнейшем только посещение врача вызывает у человека условнорефлекторное повышение частоты сердечных сокращений. Условно-рефлекторная регуляция кровообращения Условнорефлекторная регуляция сердечной деятельности Тот факт, что различные эмоции вызывают изменение сердечной деятельности, указывает на значение коры полушарий большого мозга в регуляции деятельности сердца. Доказательством этого является то, что изменения ритма и силы сердечных сокращений можно наблюдать у человека при одном упоминании или воспоминании о факторах, вызывающих у него определенные эмоции. Наиболее убедительные данные о наличии корковой регуляции деятельности сердца получены методом условного рефлекса. Если какой-нибудь, например звуковой, раздражитель сочетать многократно с надавливанием на глазное яблоко, вызывающим уменьшение частоты сердечных сокращений, то затем один этот раздражитель вызывает урежение сердечной деятельности — условный глазосердечный рефлекс. Условнорефлекторные реакции лежат в основе тех явлений, которые характеризуют так называемое предстартовое состояние спортсменов. Перед соревнованием у них наблюдаются изменения дыхания, обмена веществ, сердечной деятельности такого же характера, как и во время самого соревнования. (У конькобежцев на старте сердечная деятельность учащается на 22—35 сокращений в минуту). Кора мозга обеспечивает приспособительные реакции организма не только к текущим, но и к будущим событиям. По механизму условных рефлексов сигналы, предвещающие наступление этих событий или значительную вероятность их возникновения, могут вызвать перестройку функций сердца и всей сердечно-сосудистой системы в той мере, в какой это необходимо, чтобы обеспечить предстоящую деятельность организма. При чрезвычайно сложных ситуациях (действие «чрезвычайных раздражителей», по И. П. Павлову) возможны нарушения и срывы этих корковых высших регуляторных механизмов (неврозы по И. П. Павлову). При этом наряду с расстройствами поведенческих реакций (и невротическими изменениями психологического статуса человека) могут появиться и значительные нарушения деятельности сердца и сердечно-сосудистой системы. В некоторых случаях эти нарушения могут закрепиться по типу патологических условных рефлексов. При этом нарушения сердечной деятельности могут возникнуть при действии одних лишь условных сигналов. Конец формы 9. Промежуточные механизмы регуляции артериального давления. Роль транскапиллярногообмена. Промежуточные (по времени) регуляторные механизмы изменения транскапиллярного обмена; Увеличение артериального и/или венозного давления, как правило, сопровождается повышением давления в капиллярах. Напротив, при падении давления происходят обратные изменения: реабсорбция в капиллярах возрастает, артериальное давление повышается. РАССЛАБЛЕНИЕстенок сосудов;расслабление стенок вен при увеличении венозного кровенаполнения. ренин-ангиотензиновая систем  К 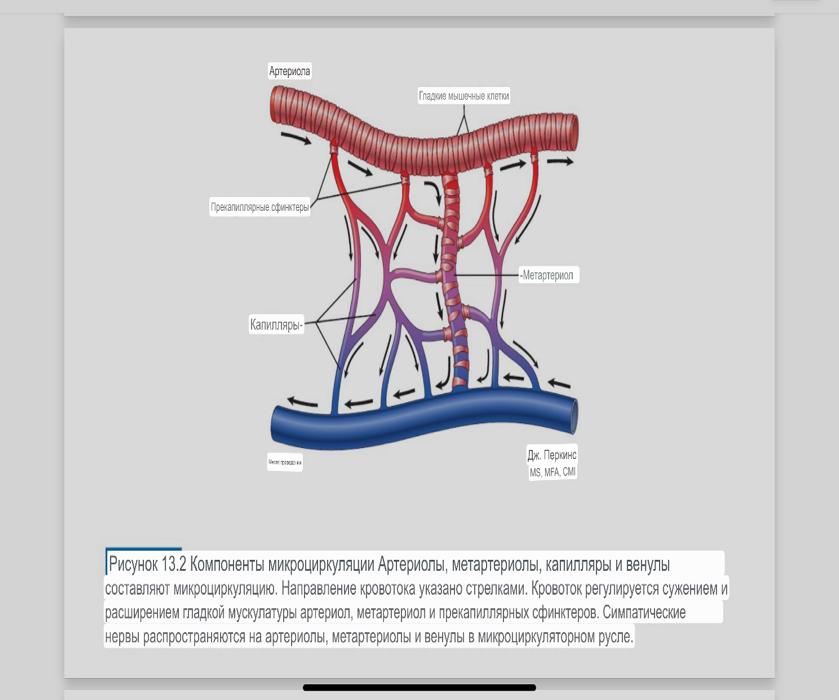 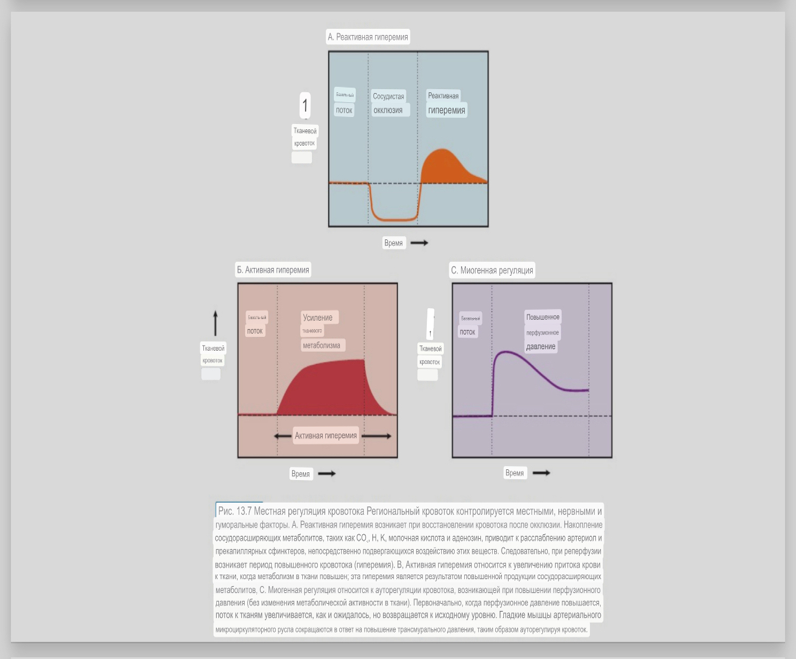 онец формы 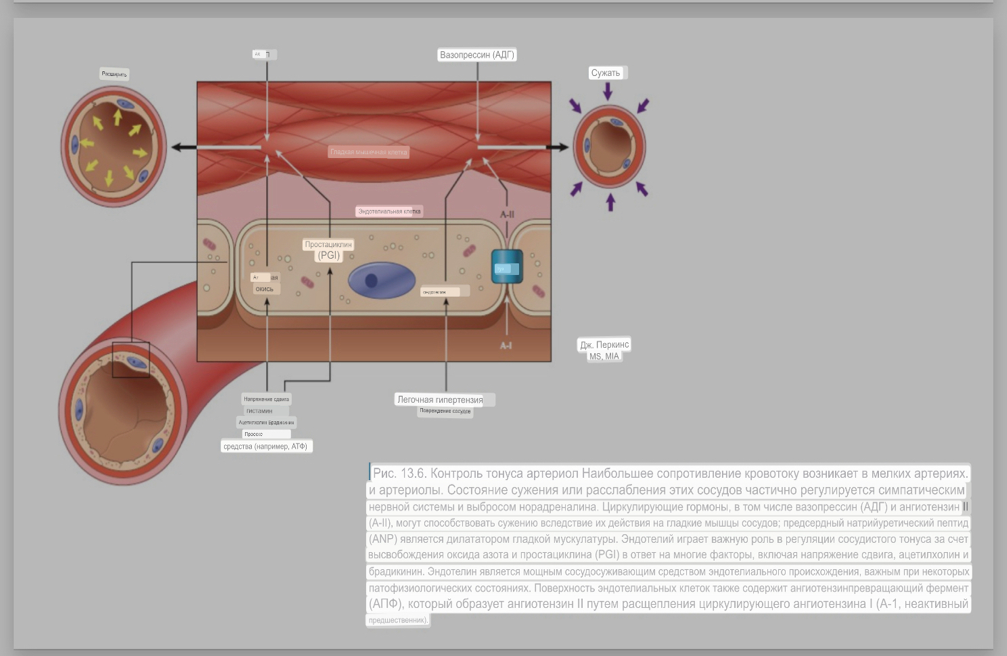 |