диплом. Аллельные варианты геновкандидатов подверженности туберкулезу у русского населения Западной Сибири
 Скачать 0.57 Mb. Скачать 0.57 Mb.
|

3. Результаты и обсуждениеУчитывая поставленные задачи, исследование включало три аспекта: изучение популяционной распространенности полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN, анализ связи исследованных генов с туберкулезом и поиск ассоциаций с патогенетически важными параметрами заболевания у русских жителей г. Томска. К настоящему времени получены результаты исследования аллельных вариантов генов подверженности к ТБ у тувинцев, выполненного по аналогичной схеме и с использованием того же набора полиморфизма генов [Рудко А.А. и др., 2003]. Это дало возможность провести сравнение полученных результатов между русскими жителями г. Томска и тувинцами. 3.1 Распространенность полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN среди здоровых лиц (контрольная группа)В настоящее время во многих популяциях мира достаточно широко исследованы полиморфные варианты гена NRAMP1, и в меньшей степени изучена распространенность аллелей генов VDR, IL12B, IL1B, IL1RN [Рудко А.А. и др., 2003; Имангулова М.М. и др., 2004; Bellamy R. et al., 1998; Ryu S. et al., 2000; Cervino A.C.L. et al., 2000;. Gao P. S., 2000; Baghdadi J. et al., 2004; Liu W. et al., 2004;]. Результаты исследований показали, что полиморфизм этих генов вносит вклад в возникновение туберкулеза. Однако известно, что восприимчивость к инфекционному заболеванию определяется одновременно многими генами с различным вкладом каждого из них в формирование того или иного патологического фенотипа. К тому же, один и тот же ген может участвовать в формировании чувствительности (или резистентности) к нескольким инфекционным заболеваниям. Вероятно, для каждого гена (и их ансамблей) существует свое "поле действия", которое модифицируется средой [Пузырев В.П., 2000]. Сочетания генов предрасположенности к болезни могут быть неодинаковы в популяциях, обусловливая различия в подверженности к заболеванию у разных народов. В связи с этим перспективным направлением исследований генетических основ предрасположенности к туберкулезу является изучение вкладов конкретных сочетаний аллелей в подверженность к болезни в различающихся как по расовой, так и по этнической принадлежности популяциях. У здоровых жителей г. Томска распределение генотипов по всем изученным полиморфным вариантам гена NRAMP1 (469+14G/C, D543N, 1465-85 G/A, 274 C/T), VDR (B/b, F/f), а также генов интерлейкинов (полиморфизм 1188A/C гена IL12B, полиморфизм +3953 A1/A2 гена IL1B) соответствовало ожидаемому при равновесии Харди-Вайнберга (РХВ), причем для большинства полиморфизмов наблюдаемая гетерозиготность (Hobs) превышала ожидаемую (Hexp) (табл.6). Лишь для частот генотипов VNTR полиморфизма гена IL1RN показано отклонение от ожидаемых при РХВ (χ2=16,75 р=0,010). При этом наблюдаемое количество гомозигот А2А2 превышало ожидаемое в 2,5 раза, а уровень гетерозиготности был меньше ожидаемого (D= -0,280). Возможно, этот факт объясняется тем, что анализируемая популяционная группа индивидов была выбрана не случайным образом из общей популяции, а включала только здоровых в отношении туберкулезной инфекции. Сравнение распространенности полиморфизма генов NRAMP1, IL12B, VDR, IL1B, IL1RN у здоровых от туберкулеза русских и тувинцев показало статистически значимые отличия между этими этническими группами, которые имели место в распределении, как частот аллелей, так и генотипов по большинству изученных генов (табл. 7). Максимальные отличия между сравниваемыми этническими группами выявлены для полиморфизма B/b гена VDR, VNTR полиморфизма гена IL1RN и 1188А/С гена IL12B. Таблица 6 Частоты аллелей и генотипов исследованных генов у здоровых жителей г. Томска
Примечание. N.O. и N.E. - наблюдаемая и ожидаемая численности генотипов соответственно; χ2 - критерий для сравнения ожидаемого и наблюдаемого распределения генотипов; d.f. - число степеней свободы; Hobs и Hexp - соответственно наблюдаемая и ожидаемая гетерозиготность; D - относительное отклонение наблюдаемой гетерозиготности от ожидаемой; * - p < 0,05 Таблица 7 Частоты аллелей и генотипов исследованных генов у русских жителей г. Томска и тувинцев
Примечание. N - численность лиц с соответствующими генотипами; р - достигнутый уровень значимости У русских г. Томска по сравнению с тувинцами аллели b гена VDR и А1 гена IL1RN встречались реже, а аллель 1188А гена IL12B - чаще. Кроме того, рассматриваемые популяционные группы статистически значимо различались по частотам аллелей и генотипов полиморфизмов 469-14G/C, D543N гена NRAMP1 и F/f гена VDR. Так, у русских чаще, чем у тувинцев наблюдали аллель 543D и реже аллель 469+14G гена NRAMP1. Индивиды, гомозиготы и гетерозиготы по аллелю f гена VDR, чаще встречались среди жителей г. Томска. Распределение генотипов полиморфизма 1465-85G/A гена NRAMP1 и +3953A1/A2 гена IL1B в исследованных популяционных группах не отличалось, однако частоты аллелей этих полиморфизмов были статистически значимо ниже у русских. Лишь для варианта 274С/Т гена NRAMP1 не найдено отличий по частотам аллелей и генотипов у русских и тувинцев. К настоящему времени накоплены результаты многочисленных исследований роли генов-кандидатов туберкулеза в патогенезе заболевания у представителей различных этнических групп. Это позволило провести сравнительный анализ частот генотипов между русскими г. Томска и другими изученными популяциями мира. Найдены статистически значимые отличия русских от других этносов по частотам аллелей генов NRAMP1, VDR, IL12B (табл. 8). Частота аллеля 469+14G NRAMP1 у жителей г. Томска оказалась статистически значимо ниже, чем у африканцев из Гамбии, но выше, чем у китайцев [Bellamy R. et al., 1998; Liu W. et al., 2004]. Самым широко исследованным полиморфизмом гена NRAMP1 оказался D543N. Во все изученных популяциях этот генный маркер был низко полиморфным. Русские отличались по частоте аллелей D543N от коренных жителей Башкирии, у которых аллель 543N вообще не обнаружен [Имангулова М.М. и др., 2004]. При сравнении с другими исследованными популяциями (китайцы, корейцы, японцы, татары, гамбийцы) статистически значимых отличий для этого полиморфного варианта не показано (рис. 2, табл. 8) [Имангулова М.М. и др., 2004; Bellamy R. et al., 1998; Ryu S. et al., 2000; Gao P. S. et al., 2000; Liu W. et al., 2004].  Рис. 2. Частоты аллелей гена NRAMP1 у русских г. Томска и в других популяциях мира (по данным литературы). * - р < 0,05 Рис. 2. Частоты аллелей гена NRAMP1 у русских г. Томска и в других популяциях мира (по данным литературы). * - р < 0,05Таблица 8 Частоты аллелей генов-кандидатов туберкулеза в различных популяциях мира в сравнении с русскими г. Томска
Примечание. χ2 - критерий использован для сравнения частот аллелей; р - достигнутый уровень значимости 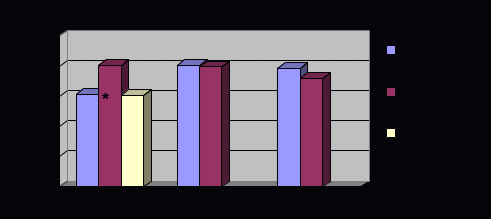 Рис. 3. Частоты аллелей полиморфизма F/f гена VDR, +3953A1/A2 гена IL1B, VNTR гена IL1RN у русских г. Томска и в других популяциях (по данным литературы). * - р < 0,05 Не показано различий при сравнении частот аллелей полиморфизма F/f гена VDR у русских г. Томска и китайцев [Liu W. et al., 2004]. Однако при сравнении с индийцами Лондона найдены статистически значимые отличия [Wilkinson R.J. et al., 2000]. В исследованной выборке русских аллель F гена VDR встречался реже (рис. 3, табл. 8). Для полиморфных вариантов +3953А1/А2 гена IL1B и VNTR гена IL1RN не выявлено отличий частот аллелей у русских и индийцев Лондона. Аллель 1188А гена IL12B у жителей г. Томска встречался чаще, чем у камерунцев (χ2 =18,37 р=0,000). Не выявлено различий в распространенности этого маркера при сравнении с англичанами, ирландцами и греками (рис. 4, табл. 8) [Hall M.A. et al., 2000]. Установленные различия частот аллелей сравниваемых генов между русским населением г. Томска и другими популяциями мира свидетельствуют об этнической специфичности генов-кандидатов подверженности к туберкулезу. Возможно, это является одной из причин дифференциальной распространенности ТБ в этнических группах. Интересными представляются выявленные отличия по частотам аллелей для всех анализируемых маркеров у русских г. Томска при сравнении с жителями Тувы. Кроме того, известно, что по частотом аллелей рассматриваемых генов тувинцы значительно отличались не только от русских, но и от представителей иных популяционных групп [Рудко А.А. и др., 2003]. Вероятно, этот факт обусловлен длительной изоляцией тувинцев от других этносов, что привело к формированию уникального генофонда [Пузырев В.П. и др., 1999]. 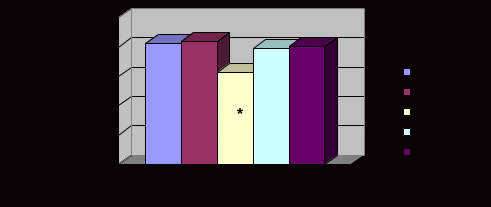 Рис. 4. Частоты аллелей полиморфизма 1188А/С гена IL12B у русских г. Томска и в других популяциях мира (по данным литературы). * - р < 0,05 Известно, что между неаллельными генами, расположенными близко на хромосоме, может возникнуть неравновесие по сцеплению в силу того, что в процессе мейоза кроссинговер между ними происходит реже, чем между далеко расположенными локусами. К тому же, под воздействием факторов популяционной динамики (естественного отбора) "сцепленными" могут оказаться гены, локализованные на разных хромосомах [Животовский Л.А., 1984]. Поэтому был проведен анализ гаметического неравновесия между исследованными полиморфными вариантами. Известно, что гены NRAMP1, IL1B и IL1RN расположены на длинном плече второй хромосомы, ген VDR - на двенадцатой, а ген IL12B - на пятой хромосоме. У русских г. Томска установлено, что в неравновесии по сцеплению находятся четыре пары полиморфизмов гена NRAMP1: 469+14G/C и 274C/T, 469+14G/C и 1465-85G/A, 274C/T и 1465-85G/A, D543N и 1465-85G/A (табл. 9). Во всех случаях неравновесия в фазе притяжения были часто встречающиеся аллели. Полиморфизм 274С/Т расположен в третьем экзоне гена, а 469+14G/C - в четвертом интроне. Такая локализация их в гене легко обьясняет наблюдаемое между ними сцепление (+0,104). Полиморфизм 1465-85G/A и D543N находятся в 13 интроне и 15 экзоне соответственно, что обусловливает неравновесие по сцеплению между ними (+0,017). Сила сцепления между полиморфизмом 1465-85G/A и вариантами 469+14G/C, 274C/T была примерно на одном уровне и составила +0,078 и +0,085, соответственно. Интересным представляется тот факт, что пары полиморфизмов гена NRAMP1, оказавшихся в неравновесии по сцеплению, были идентичны у тувинцев и русских [Рудко А.А., 2004]. Кроме того, у жителей г. Томска, в фазе притяжения оказались аллели гена VDR: b и F, сила сцепления между ними составила +0,053. Аллельный вариант F/f гена VDR у русских оказался в фазе отталкивания с полиморфизмами 469+14G/C, 274C/T, 1465-85G/A гена NRAMP1, мера неравновесия для них была -0,040, -0,039, -0,058, соответственно. Во всех случаях в фазе отталкивания оказались часто встречаемые аллели. Таблица 9 Неравновесие по сцеплению между парами исследованных полиморфных вариантов генов у русских г. Томска
Примечание. Над центральной диагональю указаны значения меры неравновесия по сцеплению (D), под диагональю - соответствующее значение χ2 для теста на неравновесие по сцеплению. Выделены величины, для которых достигнутый уровень значимости составил 5% и менееТакже в фазе отталкивания находились аллели 1465-85G гена NRAMP1 и 1188А гена IL12B, 469+14G гена NRAMP1 и +3953А1 гена IL1B, мера сцепления для них была -0,041 и -0,030. Естественный отбор в теории популяционной генетики является важнейшим фактором эволюции, вызывающим адаптивные изменения в генетической структуре популяций. В случае формирования гаметического неравновесия между маркерами под воздействием естественного отбора, определенные комбинации аллелей разных локусов дают селективные преимущества их носителям и, следовательно, сохраняются при отборе и накапливаются в популяции [Алтухов Ю.П., 2003]. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||