Коллоквиум по биологии. коллоквиум по биологии. Билет 1. Биология как комплексная наука. Современные направления в биологии. Связь биологии с другими науками. Практическое значение биологических знаний.
 Скачать 156.84 Kb. Скачать 156.84 Kb.
|

|
Билет №9. Клеточный цикл: интерфаза и деление. Митоз, значение митоза, фазы митоза. Соматические и половые клетки. Регуляция деления клеток, нарушение регуляции как причина заболеваний. Клеточный цикл (жизненный цикл клетки) — время существования клетки от начала одного деления до начала следующего деления, состоит из интерфазы и собственно процесса деления. Интерфаза — период между делениями, в котором происходят процессы роста и развития клетки, удвоения ДНК, синтеза белков и органических соединений. Деление клетки бывает прямым (амитоз) и непрямым (митоз, мейоз). Митоз — процесс непрямого деления соматических клеток эукариот, в результате которого из одной диплоидной материнской клетки образуются две дочерние с таким же набором хромосом. Выделяют четыре фазы митоза: профазу, метафазу, анафазу и телофазу. Профаза В ядре молекулы ДНК укорачиваются и скручиваются (спирализуются), образуя компактные хромосомы. Каждая хромосома состоит из двух молекул ДНК (двух хроматид), соединённых центромерой. Ядерная оболочка распадается. Хромосомы неупорядоченно располагаются в цитоплазме. Растворяются ядрышки. Начинает формироваться веретено деления, часть нитей которого прикрепляется к центромерам хромосом. В животной клетке начинают расходиться центриоли. Метафаза Хромосомы располагаются на экваторе клетки, образуя метафазную пластинку. Хроматиды соединены в области первичной перетяжки с нитями веретена деления. Центриоли располагаются у полюсов клетки. Анафаза Каждая хромосома, состоящая из двух хроматид, разделяется на две идентичные дочерние хромосомы. Дочерние хромосомы растягиваются нитями веретена деления к полюсам клетки. У каждого полюса оказывается одинаковый генетический материал. Телофаза Хромосомы раскручиваются. Вокруг хромосом начинают формироваться ядерные оболочки. В ядрах появляются ядрышки. Нити веретена деления разрушаются. Биологическое значение митоза: В результате митоза образуются генетически одинаковые дочерние клетки с тем же набором хромосом, что был у материнской клетки. Сохраняется преемственность в ряду клеточных поколений. Половые клетки (гаметы) — репродуктивные клетки, имеющие гаплоидный (n, одинарный) набор хромосом и участвующие, в частности, в половом размножении. При слиянии двух гамет в половом процессе образуется зигота, развивающаяся в особь (или группу особей) с наследственными признаками обоих родительских организмов, произведших гаметы. Соматические клетки — клетки, составляющие тело (сому) многоклеточных организмов и не принимающие участия в половом размножении. То есть это все клетки, кроме гамет. Имеют диплоидный набор хромосом (2n). Регуляция клеточного деления складывается из регуляции каждого из этих событий и организации их взаимодействия, при котором в клеточном делении устанавливается последовательность процессов и вырабатываются сигналы для инициации следующего по порядку процесса. Во-первых, деление многих видов клеток находится под контролем факторов роста, вырабатываемых другими клетками. Во-вторых, большинство нормальных клеток перестают делиться при недостатке места для новых клеток. В-третьих, многие тканевые культуры перестают расти. Последствиями нарушения клеточного цикла является потеря способности определенных клеток к делению, старения клеток, гибель клеток, возникновение опухолей. Причинами нарушений клеточного цикла могут быть недостаток питательных веществ. Примером нарушения регуляции клеточного цикла является неконтролируемый деление клеток и возникновения опухолей. . Билет №10. Клеточный метаболизм. Ферментативный характер реакций обмена веществ. Этапы энергетического обмена. Аэробное и анаэробное дыхание. Роль клеточных органоидов в процессах энергетического обмена. Пластический обмен. Автотрофы и гетеротрофы. Фотосинтез. Фазы фотосинтеза. Хемосинтез. В клетке постоянно происходит обмен веществ и энергии с окружающей средой. Обмен веществ - основное свойство живых организмов. На клеточном уровне метаболизм включает два процесса: анаболизм и катаболизм. Ассимиляция - совокупность реакций биологического синтеза. Диссимиляция - совокупность реакций расщепления веществ. Ферменты — биологические катализаторы, ускоряющие реакции обмена в клетке. Подобная локализация обеспечивает последовательность реакций. Высокая активность и специфичность действия ферментов: ускорение в сотни и тысячи раз каждым ферментом одной или группы сходных реакций. Условия действия ферментов: определенная температура, реакция среды (рН), концентрация солей. Изменение условий среды, например рН, — причина нарушения структуры фермента, снижения его активности, прекращения действия. Энергетический обмен в клетках большинства аэробных организмов в присутствии кислорода состоит из последовательных этапов: 1.подготовительного, 2.бескислородного (анаэробного), 3. кислородного (аэробного). Анаэробное дыхание - совокупность процессов, осуществляющих окисление органических веществ и получения энергии при отсутствии кислорода Аэробное дыхание - совокупность процессов, осуществляющих окисление органических веществ и получения энергии с участием кислорода. Центральным органоидом, который обеспечивает окислительно-восстановительные процессы, являются митохондрии. Функция митохондрий заключается в образовании энергии, необходимой для жизнедеятельности клеток. Источником энергии в клетке могут служить различные соединения: белки, жиры, углеводы. Пластический обмен (ассимиляция, или анаболизм) — совокупность реакций синтеза органических веществ в клетке с использованием (затратой) энергии. Пластический обмен свойственен всем живым организмам. Он необходим для обеспечения клетки строительным материалом. Так, например, в процессе фотосинтеза образуется глюкоза, а синтез белков способствует образованию новых клеток, позволяет тканям организма расти. По источнику углерода(способу питания) различают: 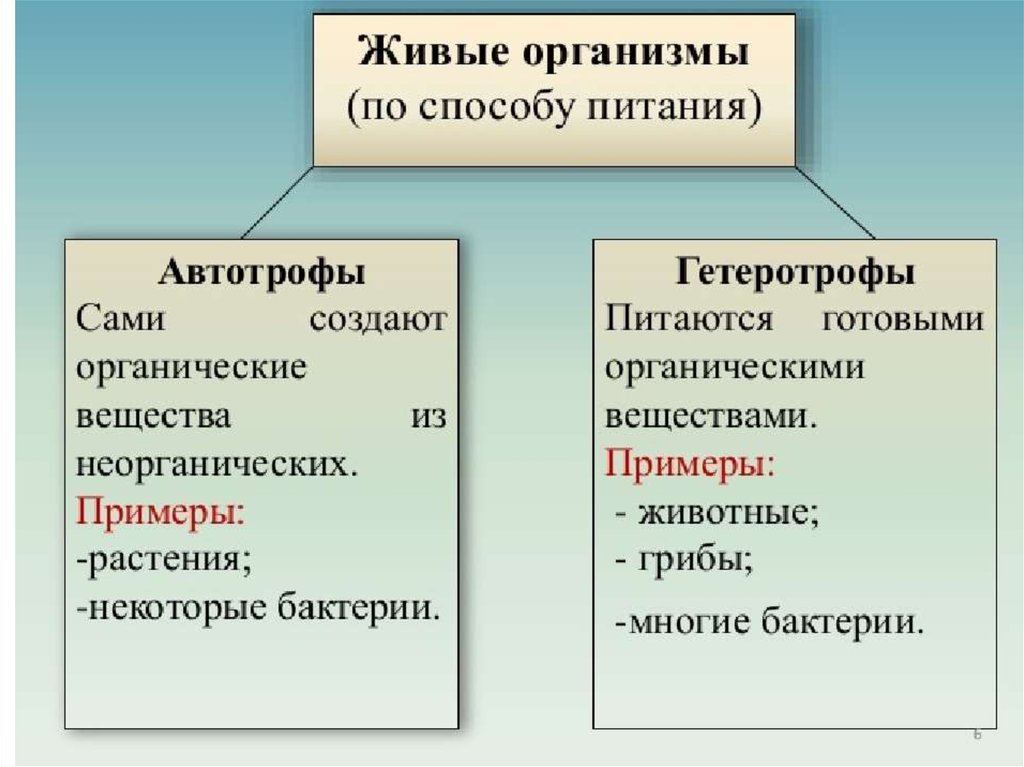 Автотрофы- осуществляют фотосинтез или хемосинтез. Фотосинтез — сложный многоступенчатый процесс; реакции фотосинтеза подразделяют на две группы: реакции световой фазы и реакции темновой фазы. Световая фаза: Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Темновая фаза: Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ. Хемосинтез: Синтез органических соединений из углекислого газа и воды, осуществляемый за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий. Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др. Билет №11. Наследственная информация и её реализация в клетке. Генетический код, его свойства. Эволюция представлений о гене. Современные представления о гене и геноме. Биосинтез белка, реакции маточного синтеза. Регуляция работы генов и процессов обмена веществ в клетке. Наследственная информация – это генетическая информация, которая передается по наследству от родителей потомству. Она обеспечивает преемственность поколений и непрерывность существования биологических видов. Реализация генетической информации Реализация генетической информации в живых клетках (то есть синтез белка, кодируемого геном) осуществляется при помощи двух матричных процессов: транскрипции (то есть синтеза мРНК на матрице ДНК) и трансляции генетического кода в аминокислотную последовательность (синтез полипептидной цепи на мРНК). Для кодирования 20 аминокислот, а также сигнала «стоп», означающего конец белковой последовательности, достаточно трёх последовательных нуклеотидов. Набор из трёх нуклеотидов называется триплетом. Принятые сокращения, соответствующие аминокислотам и кодонамгеном транскрипциимРНКДНКтрансляциимРНКтриплетомкодонам. Генетический код – это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке. Генетический код имеет несколько свойств. Триплетность. Вырожденность или избыточность. Однозначность. Полярность. Неперекрываемость. Компактность. Универсальность. Материальным субстратом гена служат молекула ДНК и в отдельных случаях РНК. Первое свидетельство такого взгляда было обнаружено при анализе явления трансформации, открытого Гриффитом в 1927 г. Однако, только в 1944 г. Эйвери и др. установили, что вещество, которое трансформирует одну форму пнемнококков в другую форму, представляет собой молекулу ДНК. Однако и после этой работы высказывались сомнение что не ДНК, а малая примесь белка ответственна за явление трансформации. Очистка препарата была доведена до того, что на 1мг. ДНК приходилась одна молекула белка. При этом, если ввести препарат ДНК - азу, разрушающий ДНК, то трансформирующая активность препарата исчезла. С другой стороны, введение ферментов, разрушающих белки, не влияло на активность препарата. Т. О., связь гена с ДНК была реально установлена в опытах в 1944г. Ген – это участок молекулы ДНК (у некоторых вирусов РНК),кодирующий первичную структуру полипептида, молекулы тРНК, рРНК, либо взаимодействующий с регуляторным белком. Ген имеет дискретную структуру. Структурная единица гена, на уровне которой осуществляются мутации и рекомбинации, является одна пара нуклеотидов - сайт (site). Количество пар нуклеотидов гена может составлять от 150 до нескольких тысяч. Самые короткие гены РНК проймазы (10 п.н.) и тРНК (70-80 п.н.) Гены эукариот, кодирующие порядок аминокислот в молекуле полипептида имеют прерывистую структуру, интроны(молчащие участки) чередуются с экзонами (смысловые). Суммарная длина интрона во много раз превышает длину экзонов. Начальная, инициирующая, и концевая, терминирующая, части гена имеют особое устройство. Ген – сложная уникальная структура, характеризующаяся специфическими особенностями в зависимости от его функций. К реакциям матричного синтеза относят репликацию ДНК, синтез и-РНК на ДНК (транскрипцию) и синтез белка на и-РНК (трансляцию), а также синтез РНК или ДНК на РНК вирусов. Биосинтез белка — это один из видов пластического обмена, в ходе которого наследственная информация, закодированная в генах ДНК, реализуется в определенную последовательность аминокислот в белковых молекулах. В биосинтезе белка выделяют два основных этапа: транскрипцию и трансляцию. Регуляция метаболизма осуществляется на двух основных уровнях – генетическом и биохимическом. На генетическом уровне обмен веществ регулируется путем регуляции экспрессии генов, а именно усилением или подавлением транскрипции и трансляции. Второй уровень регуляции – биохимический осуществляется за счёт регуляции активности ферментов. Эффекторами, способными «включать» или «выключать» гены, являются: • в катаболических генах (оперонах)– самисубстраты (например, углеводы), которые подлежат расщеплению, они выступают активаторами для белка-регулятора, т.е. выполняют функцию «включателей» гена; • в анаболических оперонах – конечные продукты синтеза (например, аминокислоты, нуклеотиды), они выступают в роли корепрессоров для белка-регулятора и, связываясь с ним, «выключают» транскрибирование ферментов, необходимых для их собственного синтеза. Механизмы регуляции метаболизма на генетическом уровне впервые были изучены на прокариотах (в оперонах кишечной палочки) в работах Жакоба и Моно еще в 40х-60х годах ХХ столетия. В настоящее время установлено, что регуляция экспрессии генов осуществляется на уровне транскрипции – при синтезе и-РНК и на уровне трансляции – при синтезе белка на рибосомах. На уровне транскрипции выявлены такие механизмы регуляции: • положительный и отрицательный контроль; • индукция и репрессия; • аутогенный контроль; • катаболитная репрессия; • смешанные механизмы регуляции; • регуляция посредством взаимодействия с энхансерами и сайленсерами (у эукариот) На уровне трансляции выявлены следующие механизмы: • аттенуация путем образования альтернативных шпилек на и-ДНК (изучена у прокариот), • регуляция трансляции на уровне сборки рибосом; • регуляция трансляции с помощью факторов инициации, элонгации и терминации. |