Курсовая работа Прилепин О.С. 2 курс ЗФО 1 подгруппа. Ботаника
 Скачать 0.69 Mb. Скачать 0.69 Mb.
|

1 2 МИНИСТЕРСТВО ПРОСВЕЩЕНИЯ РОССИЙСКОЙ ФЕДЕРАЦИИ ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ОБРАЗОВАНИЯ «ВОРОНЕЖСКИЙ ГОСУДАРСТВЕННЫЙ ПЕДАГОГИЧЕСКИЙ УНИВЕРСИТЕТ» КАФЕДРА БИОЛОГИИ РАСТЕНИЙ И ЖИВОТНЫХ ОСОБЕННОСТИ МОРФО-АНАТОМИЧЕСКОГО СТРОЕНИЯ ЛИСТА РАЗНЫХ ЭКОЛОГИЧЕСКИХ ГРУПП НА ПРИМЕРЕ КОМНАТНЫХ РАСТЕНИЙ КУРСОВАЯ РАБОТА БАКАЛАВРА по направлению подготовки 44.03.01. Педагогическое образование профиль «Биология» Дисциплина «Ботаника» Выполнил: студент заочной формы обучения 2 курса, 1 группы естественно-географического факультета Прилепин Олег Сергеевич Научный руководитель: к. б. н. доцент кафедры биологии растений и животных Дрожжина Вероника Николаевна Воронеж – 2020
1.2Общая характеристика растений разных экологических групп в зависимости от увлажнения 15 Глава 2. Результаты исследований 19 2.1 Объект исследование комнатные растения 19 2.2 Особенности анатомического строения листа мезофитов, суккулентов, гидрофитов на примере комнатных растений…………………....................... 24 Заключение 33 Список литературы 35 Введение Лист – это вегетативный орган высших растений и боковая часть побега, выполняющий функцию фотосинтеза, дыхания и транспирации. Поэтому в его структуре преобладает анатомические элементы паренхимного типа. Основной тканью листа является мезофилл, содержащий фотосинтезирующие элементы. Верхний эпидермальный слой клеток выполняет функцию газообмена и транспирации. Механическую поддержку выполняет склеренхима и колленхима. Различные экологические группы растений имеют специфическое анатомическое строение листьев, которое формирует их основные функции. Таким образом, растения способны произрастать и приспосабливаться к определенным условиям окружающей среды. Поэтому, например, для растений в засушливых условиях обитания в анатомическом строении листа будут хорошо развиваться покровные ткани – эпидермис, несущий выросты волосков. У растений, которые обитают во влажных местах, будет хорошо развита хлоренхимная ткань с большим числом устьиц. У водных растений плохо развита механическая ткань, и слабо разделенные на губчатый и столбчатый ткани мезофилла. Цель исследования: на примере комнатных растений разных экологических групп выявить особенности морфо-анатомического строения листа. Задачи исследования: 1. Изучить анатомическое строение листа Пеларгонии крупноцветковой (лат. Pelargonium grandiflorum) и фикуса Бенджамина (лат. Ficus benjamina), как представителей группы мезофитных растения; 2.Познакомиться с особенностями внутреннего строения листа суккулентного растения Алоэ древовидного (лат. Aloe arborescens Mill); 3.Выявить особенности внутреннего строения Элодеи канадской (лат. Elodea canadensis), как растения группы гидрофитов. Глава 1. Обзор литературы Общая характеристика морфо-анатомического строения листа Лист – один из вегетативных органов высших наземных растений, занимающий боковое положение на стебле. Для листа характерны следующие особенности. Он возникает экзогенно – из наружных слоёв меристемы конуса нарастания стебля в виде листового бугорка. Специфика физиологической значимости листа приводит к выразительному развитию у него ассимиляционной паренхимы (хлоренхимы). Для листа характерен ограниченный верхушечный рост. Ограниченность роста листа связана, прежде всего, с тем, что у семенных растений он почти сразу утрачивает возможность к верхушечному нарастанию и не сохраняет собственного меристематического апекса. Лист – орган моносимметричный, так как обладает лишь одной плоскостью симметрии. Для него характерна плоская форма, что делает лист бифациальным, то есть с двумя поверхностями. Плоскость симметрии перпендикулярна к двум его поверхностям – верхней (брюшной, или вентральной) и нижней (спинной, или дорсальной). Это связано с положением листового зачатка в почке, будущая верхняя сторона в таком случае будет прилегать к оси, а будущая нижняя будет обращена наружу. У однодольных растений, у которых лист ориентирован вертикально, образуются унифациальные листья с одной поверхностью. Лист характеризуется не продолжительным периодом жизни. Так, например, продолжительность жизни листа у травянистых и листопадных древесных растений несколько месяцев; у хвойных растений – от тех до десяти лет Размеры листа сравнительно разные и колеблются в пределах 2-10 см, но в определённых условиях могут достигать пару десятков метров [2, 8, 15]. Морфология и классификация листьев В структуру взрослого листа входят следующие части: пластинка или несколько пластинок (у сложных листьев), черешок и основание. Также весьма часто при основании встречаются прилистники. Пластинка – расширенная плоская, наиболее важная часть типичного листа, так как именно здесь происходят главные процессы зелённого растения – фотосинтез, транспирация и дыхание. У большинства растений между основанием листа и пластинкой развивается черешок. Черешок – узкая стеблевидная часть, располагающаяся между пластинкой и основанием. Он необходим для ориентировки листа по отношению к свету. Листья неоснащённые черешком называют сидячими, а с черешками – черешчатыми. Основание – базальная часть листа, сочлененная со стеблем. Имеет вид небольшого утолщения (листовая подушечка) или же вовсе не заметно. У большинства растений оно может значительно разрастаться в длину и ширину, тем самым целиком охватывая междоузлие. Такое разросшееся основание называют листовым влагалищем. Оно особенно характерно для класса однодольных (семейства Лилейные Осоковые, Мятликовые) и двудольных (семейство Сельдерейные). Главная функция влагалища защита пазушных почек и молодых, продолжительно растущих основании междоузлий, где сохраняется вставочная меристема. Не редко у листьев встречаются прилистники. Прилистники – это парные боковые выросты основания листа. Форма и размеры прилистников, также, как и их функции – не одинаковы и варьируются в зависимости от вида растений. Как правило, они развиваются раньше, чем пластинка, предохраняя листья в почке (яблоня, берёза, липа), а при раскрывании почки они сбрасываются. Иногда, в случае зеленой окраски, они функционируют как фотосинтезирующие органы (горох, фиалка трехцветная). Прилистники бывают свободными и « приросшими » к черешку; они могут смещаться на внутреннюю сторону листа (пазушные) [1, 20]. По существующей классификации листья делят на два типа простые и сложные. Если лист состоит из одной пластинки, то его называют простым. У сложных листьев имеется несколько листовых пластинок, каждая из которых имеет свой черешок, сидящий на общей оси – рахисе. В зависимости от расположения листочков различают перисто - и пальчатосложные листья. В случае если рахис завершается на верхушке непарным листочком, лист называется непарноперистосложным, при его отсутствии – парноперистосложным. Для сложного листа так же характерно ветвление, в результате которого образуются листья дважды - и триждыперистосложные, многократно тройчатые. У пальчатосложных и тройчатосложных листьев рахиса нет. При характеристике листовой пластинки принимают во внимание целый ряд признаков: общие очертания листа, форму основания и верхушки, форму края, жилкование, характер поверхности и многие другие признаки. Форма листьев несет не только наследственный признак того или иного вида растения или целой систематической группы, но и отражает связь с условиями обитания, характерными для вида и группы. Листовая пластинка может быть цельной или расчлененной. В зависимости от глубины расчленения выделяют лопастные, раздельные и рассеченные простые листья или листочки сложного листа. При этом надрезы у лопастного листа называют лопастями, у раздельного – долями, у рассеченного – сегментами. Большая изрезанность листовой пластинки предохраняет лист, во время каких либо сильных внешних воздействий например, от разрыва ветром и дождем при слабо развитой механической ткани [1, 17, 20]. Жилкование у растений – это система проводящих пучков (жилок) в листовых пластинках, через которые осуществляется транспорт веществ. Жилки выполняют проводящую и механическую функции листа. По ксилеме лист поступают вода и минеральные соли; по флоэме из листа оттекают выработанные в нем органические вещества. Также жилки создают опору для паренхимы и предохраняют листовые пластинки от разрывов. Через основание и черешок в центре листа проходит главная жилка, от которой берут начало боковые жилки второго и последующих порядков. Между собой они также сплетаются сетью боле мелких жилок – анастомозов [11, 16, 25]. У растений выделяю следующие типы жилкования: – дихотомическое жилкование, которое свойственно большинству папоротниковидных, а также голосеменным – гинкго. При этом анастомозы отсутствуют, а окончания жилок подходят к краю листовой пластинки. – параллельное и дуговое жилкование характерно для однодольных. От основания листа в пластинку входит несколько жилок одинакового размера, пронизывая пластинку параллельно (злаки) или дугообразно (ландыш, тюльпан). По всей длине продольные жилки соединяются перемычками – анастомозами, в косом или поперечном направлении. Это функционально выгодно при проведении веществ в период частичных разрывов пластинки листа: – сетчатое, перистое или пальчатое жилкование наиболее характерно для двудольных растений. При сетчатом жилковании более мелкие разветвления жилок всегда соединяются друг с другом перемычками. При перистом жилковании из стебля в лист идёт лишь одна главная жилка, которая в дальнейшем, в пластинке распадается на сеть более мелких. При пальчатом – из черешка выходит несколько равных жилок, и каждая из которых разветвляется [11, 23]. Жилкование – несет в себе важный признак в систематике растений, так как оно дает возможность судить о том, на какой ступени эволюции находится растение [1]. Анатомия листа Важнейшей тканью листа как основного фотосинтезирующего органа растения является хлорофиллоносная фотосинтезирующая паренхима – мезофилл, занимающая его основной объем. Покрывающая поверхность листа эпидерма защищает лист от излишней потери воды и осуществляет процессы газообмена и транспирации. Механические ткани (колленхима и склеренхима) придают пластинке листа необходимую прочность и позволяют листу занимать определенное положение в пространстве. Входящие в состав проводящих пучков (жилок) ксилема и флоэма обеспечивают поступление к клеткам мезофилла воды, необходимой для фотосинтеза, и отток ассимилятов (органических веществ, образованных мезофиллом) из листа. Как правило, все ткани листа – первичные. Образование всех этих тканей проходит в ходе морфогенеза листа – процесса его формообразования. Каждый лист побега начинает развиваться из листового бугорка, формирующегося на конусе нарастания побега в зоне инициального кольца. Листовой бугорок, состоящий из активно делящихся меристематических клеток, разрастаясь, превращается в зачаток листа – листовой примордий. О 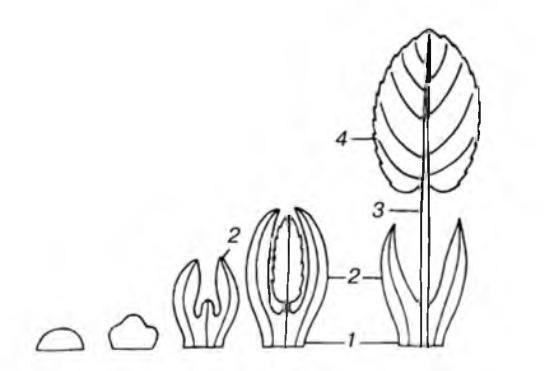 дновременно закладывается тяж прокамбия, переходящий из листового примордия в формирующуюся стеблевую часть побега. Боковые тяжи прокамбия возникают из меристематических клеток, находящихся в средней зоне зачатка листа. По мере развития листа из тяжей прокамбия образуются проводящие пучки листа, а из остальных зон меристемы формируются эпидерма, механические ткани и ассимиляционная паренхима. Размеры, форма и характер рассеченности пластинки листа определяются особенностями деления клеток краевой, или маргинальной, меристемы, из которой она формируется. Маргинальная меристема располагается в виде двух валиков вдоль осевой части листового примордия. Если ее клетки делятся равномерно, то формируется лист с цельной пластиной, если нет – с лопастной, раздельной или рассеченной [4, 5, 13]. дновременно закладывается тяж прокамбия, переходящий из листового примордия в формирующуюся стеблевую часть побега. Боковые тяжи прокамбия возникают из меристематических клеток, находящихся в средней зоне зачатка листа. По мере развития листа из тяжей прокамбия образуются проводящие пучки листа, а из остальных зон меристемы формируются эпидерма, механические ткани и ассимиляционная паренхима. Размеры, форма и характер рассеченности пластинки листа определяются особенностями деления клеток краевой, или маргинальной, меристемы, из которой она формируется. Маргинальная меристема располагается в виде двух валиков вдоль осевой части листового примордия. Если ее клетки делятся равномерно, то формируется лист с цельной пластиной, если нет – с лопастной, раздельной или рассеченной [4, 5, 13].Рис. 1. Морфогенез листа: 1– основание листа, 2 – прилистники, 3 – черешок. 4 – листовая пластинка Эпидерма покрывает лист в течение всей его жизни. Между собой клетки плотно соединяются. Такая плотность эпидермы характеризует ее роль в защите от потери излишней воды, а также как механическую опору. Эпидерма, располагающаяся на верхней стороне листа, отличается сравнительно большими клетками, чем эпидерма, находящаяся на его нижней стороне. Верхняя эпидерма, в большей степени подвергающаяся воздействию солнечных лучей, обычно покрыта и более толстой кутикулой и отличается более густым опушением. Как правило, в клетках данной ткани отсутствуют хлоропласты, но они присутствуют в замыкающих клетках устьиц. У листьев с горизонтально расположенной пластинкой устьица находятся, как правило, лишь на нижней стороне листа. При вертикальном положении пластинки устьица распределены равномерно по обеим ее сторонам. У плавающих листьев водных растений устьица образуются лишь у верхней эпидермы. Строение эпидермы может сильно варьироваться в зависимости от местообитания растений [13, 26]. Клетки эпидермы также способны образовывать некие выросты – трихомы, которые бывают разнообразными (волоски, сосочки, щетинки). Их образование приурочено к ранним этапам развития листа. Главная роль этих выростов – защитная (от перегрева солнцем, от потери воды).   Рис. 2. Объемное изображение фрагмент пластинки дорсо - вентрального листа (по Т. И. Серебряковой и др., 2006) Мезофилл занимает все пространство между нижней и верхней эпидермой, исключая проводящие и механические (арматурные) ткани. Клетки мезофилла относительно схожи по форме, в большинстве своем округлые или слегка вытянутые, также возможны выросты [6]. Протопласт состоит из постепенного слоя цитоплазмы с ядром и многочисленными хлоропластами. Внутри клетки содержится центральная вакуоль. Мезофилл в обычном случае дифференцирован на два вида ткани: столбчатую (палисадную) и губчатую. Клетки палисадного мезофилла, который располагается под верхней эпидермой, вытянуты перпендикулярно поверхности листа и образуют несколько слоев. Клетки губчатого мезофилла располагаются свободней относительно друг друга, а, следовательно, межклетные пространства в этой ткани сравнительно большие по отношению к объему самих клеток. Увеличение межклетных пространств достигается различными путями: в одних случаях клетки сохраняют округлую форму, а в других образуют выросты. Палисадная ткань содержит примерно 3/4 - 4/5 всех хлоропластов листа выполняет главную работу по ассимиляции углекислого газа. Именно поэтому палисадная ткань имеет расположение в наиболее выгодных условиях освещения, непосредственно под верхней эпидермой. Благодаря тому, что клетки вытянуты перпендикулярно поверхности листа, лучи света проще проникают вглубь мезофилла, встречая на пути меньше препятствий. Через губчатый мезофилл происходит газообмен. Углекислый газ, рассеянный в атмосфере, через устьица, расположенные главным образом с нижней стороны эпидермы, проникает в межклетники губчатого мезофилла и беспрепятственно распределяется внутри листа. Кислород, выработанный мезофиллом при фотосинтезе, перемещается в обратном направлении и через устьица выходит наружу, в окружающую атмосферу. В темное время суток, когда процесс фотосинтеза прекращается, но продолжается, дыхание, направление движения газов меняется. Количество хлоропластов в его клетках в 2-6 раз меньше, чем в клетках столбчатого мезофилла, что объясняет более светлую окраску нижней стороны листа. Необходимо отметить, то, что между клетками палисадного мезофилла так же имеются межклетники, но меньшего размера, чем в губчатом. Расположение устьиц преимущественно на нижней стороне листа объясняется не только положением губчатого мезофилла. Потеря воды листом в процессе транспирации идет медленнее через устьица, расположенные в нижней, а не в верхней эпидерме. Кроме того, это выгодно для поглощения почвенного углекислого газа, который выделяется в процессе жизнедеятельности живых существ [6, 21]. Проводящая система в листьях представлена жилками, которые многократно ветвятся в листовой пластинке. В жилке может быть один или несколько проводящих пучков. Большинство пучков закрытые, лишь те, что больше – открытые, но камбий развит слабо. В черешке может находиться несколько коллатеральных пучков, расположенных в виде кольца (платан), или одиночные разных типов: коллатеральный (камелия, бересклет), биколлатеральный (тыква), концентрический (цитрусовые) [13]. проводящих пучках листовой пластинки ксилема обращена к верхней поверхности листа, а флоэма – к нижней стороне листа. В пучках, за исключением самых мелких, имеются трахеи, а во флоэме – ситовидные трубки. В мелких пучках трахеи сменяются трахеидами, во флоэме – паренхимными клетками. Окончания жилок листа состоят обычно из простых пучков. Проводящие пучки обособлены от клеток мезофилла. Обычно они окружены окаймляющей паренхимой, к которой иногда может присоединяться склеренхима. Крупные проводящие пучки погружены в паренхиму и отделены от мезофилла, а мелкие проводящие пучки пронизывают мезофилл и слепо там заканчиваются [1]. Механические ткани особенно значимы для листьев с плоскими пластинками – пронизывая все ткани пластинки, они придают ей необходимую прочность и позволяют противостоять факторам внешней среды (ветер, дождь, град и т.д.). Колленхима типично расположена в черешке, не мешая ему благодаря своей пластичности изменять положение в пространстве. В листовой пластинке она встречается только у двудольных растений – располагается под верхней эпидермой над самыми крупными жилками. У некоторых растений (фикус) она может образовываться и поджилками. Склеренхима тоже обычно приурочена к пучкам – она огибает флоэму. В некоторых случаях склеренхима может развиваться и над ксилемой. У однодольных (особенно злаков) она может окружать весь пучок. С возрастанием порядка ветвления жилок количество механических тканей вокруг них уменьшается. Вокруг мелких жилок механических тканей нет. В листьях однодольных растений склеренхима образуется не только в виде обкладки вокруг пучков, но и в виде обособленных тяжей, например, у пальм. У обитателя засушливых степей – ковыля склеренхимные обкладки пучков и связанные с ними вертикальные тяжи склеренхимы образуют своеобразные балки, пронизывающие толщу пластинки листа и придающие ей жесткость и прочность. Более того, именно наличием этих склеренхимных балок объясняют сегодня способность листа ковыля скручиваться в трубку при дефиците влаги. У некоторых растений (ирис, синеголовник) тяжи склеренхимы могут располагаться под эпидермой по краю листовой пластинки. В тканях листа в виде идиобластов встречаются и отдельные склереиды. Размеры и форма их разнообразны. В мезофилле листьев камелии и чая можно обнаружить многолучевые астросклереиды, названные так из-за звездообразной формы. У растений с зубчатым или пильчатым краем листовой пластинки (земляника, лапчатка и др.) склереиды располагаются в глубине вырезов, снижая риск ее разрыва [13, 22]. Функции и метаморфозы листа Главные функции листа фотосинтез, газообмен и транспирация. В процессе фотосинтеза в листьях образуются из воды и диоксида углерода органические вещества, главным образом сахара. Побочный продукт фотосинтеза – кислород выделяется в атмосферу. Сахара в виде раствора переносятся по проводящим тканям растения ко всем растущим органам, где служат источниками энергии и строительного15 материала. Интенсивность фотосинтеза – основа продуктивности сельскохозяйственных растений. Чем лучше будет развита листовая поверхность растения, чем меньше она будет повреждена болезнями и вредителями, тем более высокий урожай можно получить. Транспирация – регулируемое испарение, обеспечивающее подъем воды с растворенными минеральными веществами из корневой системы в листья. Кроме этого транспирация обеспечивает охлаждение растения на 5-7 °С по сравнению с температурой окружающего воздуха, что создает оптимальные условия для процесса фотосинтеза (при слишком высокой температуре он прекращается). Транспирация идет очень активно – например, одна береза испаряет в день до 200 л воды. Газообмен обеспечивает фотосинтез и дыхание растений. При дыхании растение поглощает кислород, а выделяет в атмосферу диоксид углерода. Для осуществления фотосинтеза растение поглощает из воздуха диоксид углерода, а выделяет в него кислород. Фотосинтезирующие растения – основные поставщики кислорода в атмосферу нашей планеты. Кроме основных, названных выше, листьям свойственны и другие функции. Видоизменяясь в чешуи, они могут стать местом отложения запасных веществ (луковицы) или защищать зимующие почки от иссушающего действия мороза. С помощью листьев, превратившихся в колючки, растения защищаются от травоядных животных. Усики листового происхождения, прикрепляя побеги к опорам, обеспечивают им нужное расположение в пространстве. Для многих растений характерны метаморфозы листьев. Можно выделить следующие типы метаморфозов листа: 1.Усики – нитевидные органы, весьма чувствительны к прикосновению и приспособленные для лазания. Такие растения выполняют двойную функцию – фотосинтез и захват опоры. Для некоторых видов растений характерны на концах усиков специальные липкие дисковидные вздутия – присоски. 2.колючки свойственны растениям, которые обитают в сухом и жарком климате, хотя нередко могут встречаться и в других климатических зонах. Они выполняют две основные функции: уменьшаю испаряющую поверхность надземной части растения и защищают стебли, стволы и листья от поедания животными. 3.Филлодий – это метаморфоз черешка или основания листа в образование, подобное плоской листовой пластинке, выполняющей функции фотосинтеза. 4.Ловчие аппараты насекомоядных растений. У некоторых видов покрытосеменных все листья или часть из них метаморфизированы в ловчие аппараты. Растения этих видов автотрофные, но наряду с этим при помощи ловчих аппаратов они используют богатую азотом и фосфором органическую пищу, переваривая животных [1, 13]. 1.2Общая характеристика растений разных экологических групп в зависимости от увлажнения На внешний облик растений, а, следовательно, на особенности строения листьев влияет и степень увлажнения среды обитания, в связи с этим различают четыре экологические группы растений: гидрофиты, гигрофиты, мезофиты, ксерофиты. Гидрофиты–растения,частично или полностью произрастающие вводе, у них большие тонкие листья. В воде они часто покрыты слизью. В листьях, как и в других органах, у них развита воздухо-запасающая паренхима – аэренхима. Эпидермис и кутикула слабо развиты, у листьев, находящихся в воде, нет устьиц, а у плавающих листьев они расположены на верхней поверхности листа. У листьев, погруженных в воду, мезофилл слабо дифференцирован [9, 11]. Гигрофиты–растения,произрастающие в условиях сильногоувлажнения почвы и воздуха (берега рек, озер). Листья у них широкие, крупные, мягкие с хорошо развитой хлоренхимой. Растения этой группы имеют, как правило, крупные, тонкие, мягкие листовые пластинки с хорошо развитой хлоренхимой и с небольшим числом устьиц, нередко расположенных с обеих сторон. Устьица большей частью широко открыты, в связи с этим транспирация мало отличается от физического испарения. Эпидермис почти не имеет кутикулы. Поэтому сорванные растения этой группы быстро вянут. К гигрофитам в первую очередь относят тропические растения, которые живут при высокой температуре и влажности воздуха. Нередко гигрофиты обитают в тени под пологие леса (например, папоротники) или на открытых пространствах, но обязательно на почвах, переувлажненных или покрытых водой. В умеренном и холодном климате типичными гигрофитами являются теневые травянистые растения лесов. На открытых местах и влажных почвах растут световые гигрофиты. Это такие, как калужница, плакун-трава, росянка, многие злаки и осоки сырых местообитаний, из культурных растений к световым гигрофитам относят рис, культивируемый на полях, залитых водой. Мезофиты–это растения умеренно увлажненных местообитаний.Листья имеют отличительные размеры. Листья у них большие, мягкие, нетолстые, плоские, с умеренно развитыми покровной, проводящей, механической, столбчатой и губчатой тканями. Устьица располагаются на нижней стороне листовых пластинок. Хорошо выражена регуляция устьичной транспирации. К мезофитам относятся многие луговые травы (клевер луговой, тимофеевка, ежа сборная) и большинство лесных растений (ландыш, зеленчук и др.). Значительная часть мезофитов среди лиственных деревьев (береза, липа), многих полевых (картофель) и плодово-ягодных (яблоня, вишня) культур [10, 24]. Один и тот же мезофильный вид, попадая в разные по водоснабжению условия, обнаруживает известную пластичность, приобретая во влажных условиях более гигроморфные, а в сухих – более ксероморфные черты. Мезофиты связаны переходами с другими экологическими типами растений по отношению к воде, поэтому четкую границу между ними провести зачастую очень трудно. Ксерофиты–растения засушливого климата(степи,пустыни,полупустыни) или засушливых мест обитания в умеренных широтах (очиток едкий, молодило и др.). Эти растения хорошо переносят недостаток влаги. Среди ксерофитов выделяют суккуленты и склерофиты. Неблагоприятный водный режим растений в сухих местообитаниях обусловлен ограниченным поступлением воды при ее недостатке в почве и увеличением расхода влаги на транспирацию при большой сухости воздуха и высокой температуре. Таким образом, для преодоления недостатка влаги могут быть разные пути: увеличение ее поглощения и сокращение расхода, а также способность переносить большие потери воды. При этом различают два основных способа преодоления засухи: возможность противостоять иссушению тканей, или активное регулирование водного баланса, и способность выносить сильное иссушение. Важное значение для ксерофитов имеют разнообразные структурные приспособления к условиям недостатка влаги. Наземные органы ксерофитов отличаются своеобразными чертами, носящими отпечаток трудных условий водоснабжения. У них сильно развита водопроводящая система, хорошо заметная по густоте сети жилок в листьях, подводящих воду к тканям. Эта особенность облегчает ксерофитам осуществлять пополнение запасов влаги, расходуемой на транспирацию. Структурные приспособления защитного характера у ксерофитов, направленные на уменьшение расхода воды, можно свести к следующему: 1.Общее сокращение транспирирующей поверхности за счет мелких узких, сильно редуцированных листовых пластинок. 2.Уменьшение листовой поверхности в наиболее жаркие и сухие периоды вегетационного сезона. 3.Защита листьев от больших потерь влаги на транспирацию благодаря развитию мощных покровных тканей – толстостенного или многослойного эпидермиса, нередко несущего различные выросты и волоски, которые образуют густое «войлочное» опушение поверхности листа. 4.Усиленное развитие механической ткани, предупреждающее обвисание листовых пластинок при больших потерях воды [24]. Суккуленты–многолетние растения с сочными мясистыми листьямиили стеблями. Обладают способностью накапливать в своих тканях большое количество воды. Водозапасающие ткани у них могут быть развиты в стеблях или листьях, поэтому они подразделяются на стеблевые суккуленты и листовые суккуленты.У стеблевых суккулентов листья превращены вколючки и чешуйки (кактусы, молочаи). У листовых суккулентов листья с толстым кутинизированным эпидермисом, покрытым восковым налетом (алоэ, агава, молодило). Ассимилирующие ткани представлены у них в основном губчатой паренхимой. Остальное пространство занято водозапасающей паренхимой. Устьиц на поверхности тела почти нет. А если есть, то они мелкие, располагаются в ямках и большую часть времени закрыты. Открываются только на ночь. Все это предельно сокращает транспирацию. Характерной особенностью суккулентов является высокая поглощающая способность. В период дождей отдельные виды поглощают в себя большое количество воды. Накопленную влагу суккуленты в дальнейшем медленно расходуют. Суккуленты произрастают в районах сжарким сухим климатом. Там, где хотя бы изредка проходят дожди кратковременные, но обильные, ливневые Склерофиты(от греч. «склерос»–твердый,жесткий)–растения,повнешнему виду резко отличающиеся от суккулентов. Они внешне суховатые. Листья твердые, жесткие, иногда видоизменяются в чешуйки или колючки; некоторые опушены и могут скручиваться в трубку. Склерофиты не накапливают в себе влагу, а испаряют ее в большом количестве, постоянно доставая из глубоких слоев почвы. При продолжительном прекращении подачи воды может наблюдаться сбрасывание листьев или части побегов, что приводит к сокращению испарения. Многие из ксерофитов переносят засушливое время года в состоянии вынужденного покоя [19]. 1 2 | |||||||||||||||