Структура и функции клеточного ядра. Центральная догма молекулярной биологии
 Скачать 0.84 Mb. Скачать 0.84 Mb.
|

|
Структура и функции клеточного ядра Центральная догма молекулярной биологии 1 Привести схему строения и охарактеризовать состав молекулы нуклеотида. Через какие связи нуклеотиды соединяются в полинуклеотидную цепь? Нуклеотид состоит из пуринового или пиримидинового азотистого основания, пентозы (сахар с 5 атомами углерода) и остатка фосфорной кислоты. Нуклеотиды соединены между собой ковалентно в длинные полинуклеотидные цепи. Эти цепи в подавляющем большинстве случаев (кроме некоторых вирусов, обладающих одноцепочечными ДНК-геномами) попарно объединяются при помощи водородных связей во вторичную структуру, получившую название двойной спирали.  2 Дать сравнительную характеристику строения молекул ДНК и РНК. Какие связи формируют двойную спираль ДНК? Объяснить принцип комплиментарности в построении двойной спирали, назвать комплиментарные пары нуклеотидов. Существует два вида нуклеиновых кислот – РНК (рибонуклеиновая кислота) (рис. 12) и ДНК (дезоксирибонуклеиновая кислота) (рис. 13). Состав нуклеотидов РНК и ДНК отличается: в первом случае сахар-пентоза представлен рибозой,а во втором случае дезоксирибозой (отсюда и названия соответствующих нуклеиновых кислот). Также различается и набор азотистых оснований. В РНК присутствуют аденин, гуанин, цитозин и урацил, в ДНК – аденин, гуанин, цитозин и тимин (вместо урацила). Молекула РНК представляет собой одноцепочечную молекулу, в то время как ДНК существует в виде двойной спирали – две полинуклеотидные цепочки закручены друг вокруг друга и связаны между собой водородными связями по всей длине. При этом сахаро- фосфатные остовы цепочек обращены наружу, а азотистые основания спрятаны внутри дуплекса. Таким образом, именно между азотистыми основаниями, находящимися друг против друга в противоположных цепях ДНК, и возникают водородные связи (рис. 13). При этом выполняется принцип комплементарности (дополнительности) – к образованию пар способны только комплементарные основания: аденин соединяется строго с тимином (между ними формируются две водородные связи), а гуанин – с цитозином (образуют три водородные связи). Как видим, пуриновое основание всегда соединяется с пиримидиновым, что обеспечивает постоянный диаметр спирали по всей длине ДНК. Последовательность нуклеотидов в ДНК строго уникальна для каждой молекулы и формирует первичную структуру цепочки нуклеиновой кислоты. Вторичная структура ДНК – это двойная спираль. ДНК отвечает за хранение и передачу в поколениях наследственной генетической информации, а РНК принимает непосредственное участие в реализации этой информации. 3 Дать определение понятия "транскрипция", объяснить молекулярный механизм транскрипции: что является матрицей, какой используется фермент, откуда берутся предшественники для синтеза? Синтез белка начинается с транскрипции (от лат. transcriptio – переписывание) – «переписывания» информации с ДНК на РНК, в результате чего в клетке происходит многократное копирование нужного гена. У прокариот этот процесс происходит прямо в цитоплазме; в эукариотической клетке для хранения генетического материала (ДНК) имеется специальный компартмент – клеточное ядро, там же и происходит транскрипция. При этом двухцепочечная молекула ДНК на определенном участке временно раскручивается за счет разрыва водородных связей между комплементарными нуклеотидами, а точнее, между их азотистыми основаниями. С помощью фермента РНК-полимеразы вдоль одной из цепей ДНК производится подбор свободных комплементарных нуклеотидов из ядерного сока и присоединение их по месту обнажившихся водородных связей. Таким образом, из свободных нуклеотидов бок о бок с цепью ДНК по принципу комплементарности формируется одноцепочечная молекула матричной, или информационной, РНК (мРНК, или иРНК), которая является полной копией одной из цепей ДНК, за исключением того, что вместо тимина в состав РНК по тем же позициям включается урацил (рис. 20). В этом случае транскрибируемый участок молекулы ДНК выступает в роли некоего образца, матрицы для синтеза другой макромолекулы – РНК, а в дальнейшем и белка. Такие синтезы принято называть матричными. 4 Дать определение понятия "трансляция". Привести схему и объяснить механизм работы рибосом. Определить роль каждой формы РНК в синтезе белка. Следующий этап синтеза белка называется трансляцией (от лат. translatio – перевод). В этом процессе задействованы следующие компоненты: мРНК (в качестве рабочей копии гена), рибосомы и молекулы транспортных РНК (тРНК). Рибосомы – это очень мелкие клеточные органоиды (размером всего 25 нм), не имеющие мембранного строения, предназначенные для синтеза белковых молекул. Рибосома состоит из двух субъединиц – большой и малой. Каждая субъединица представляет собой рибонуклеопротеидный комплекс, т.е. состоит из рибосомной РНК (рРНК) и определенного набора белков. Всего в состав эукариотической рибосомы входят около 80 белков (30 в малую субъединицу и 50 – в большую) и 4 молекулы РНК: 18S РНК (формирует скелет малой субъединицы), 28S РНК, 5,8S РНК и 5S РНК (образуют основу большой субъединицы). Эукариотическая рибосома в собранном состоянии имеет коэффициент седиментации 80S, а ее субъединицы – 60S и 40S. У прокариот соответствующие характеристики составляют 70S, 50S и 30S. Большая и малая субъединицы соединяются только в момент синтеза белковой молекулы; после окончания трансляции рибосома вновь разбирается на отдельные субъединицы и в таком разобранном виде существует до начала следующего своего цикла в синтезе белка. Транспортная РНК (тРНК) – это молекула особой конфигурации в виде клеверного листа (рис. 22). Такую трехлопастную конформацию молекуле тРНК придают водородные связи между комплементарными нуклеотидами, стоящими в определенных положениях вдоль линейной молекулы, стягивая соответствующие участки цепи в петли. Таким образом, тРНК сочетает в себе одно- и двухцепочечную структуры. Триплет, располагающийся на средней лопасти клеверного листа, – антикодон – предназначен для комплементарного связывания с соответствующим триплетом на мРНК – кодоном. Каждая разновидность тРНК имеет свой вариант антикодона. Напомним, что на основе 4 нуклеотидов, взятых по три штуки, теоретически можно построить 64 комбинации триплетов. Реально используется 61 комбинация, и, соответственно, существует 61 вид тРНК (см. рис. 15). Противоположный антикодону (свободный) конец молекулы тРНК предназначен для связывания со строго определенной, одной из 20, аминокислот. В момент трансляции рибосома нанизывается на нить мРНК, захватывая участок молекулы длиной в несколько триплетов нуклеотидов. При этом две субъединицы одной рибосомы соединяются, и нить матричной РНК оказывается втиснутой в щель между ними. Сюда же подходят две молекулы тРНК, нагруженные аминокислотами – каждая своей аминокислотой, если трансляция только началась, и белковой цепи как таковой еще нет. Если же процесс трансляции продвинулся хотя бы на один шаг, то одна из тРНК теперь несет уже частично синтезированную цепь белка из двух или более аминокислот. Одновременное нахождение в рибосоме молекулы мРНК и двух молекул тРНК возможно благодаря тому, что в рибосоме имеется функциональный центр с тремя различными участками для связывания РНК – один для мРНК и два для тРНК. В центр попадает участок молекулы мРНК протяженностью в два триплета (два кодона). Что касается участков для связывания тРНК, то один из них удерживает молекулу тРНК, присоединенную к растущему концу полипептидной цепи (поэтому его называют пептидил-тРНК-связывающим участком, или П-участком), а второй служит для удержания только что прибывшей молекулы тРНК, нагруженной аминокислотой, и называется аминоацил-тРНК-связывающим участком, или А- участком. К обоим участкам молекулы тРНК прикрепляются лишь в том случае, если их антикодоны оказываются комплементарными кодонам мРНК, находящимся на данный момент в рибосоме. Когда две тРНК оказываются в рибосоме рядом друг с другом, между последней аминокислотой растущей белковой цепи и вновь прибывшей аминокислотой возникает ковалентная пептидная связь – белковая цепь удлиняется еще на одну аминокислоту и «перебрасывается» с одной тРНК на другую. «Опустевшая» тРНК теряет связь с кодоном мРНК и выводится из П-участка рибосомы. Затем рибосома перемещается вдоль цепи мРНК на один триплет, при этом оставшаяся в рибосоме тРНК занимает освободившийся П-участок, а на ее место (в освободившийся А-участок) подбирается новая тРНК с очередной аминокислотой. Так рибосома движется вдоль молекулы мРНК, захватывая все новые и новые кодоны и вовлекая в процесс все новые и новые тРНК с пришитыми к ним аминокислотами. В результате формируется полипептид с уникальной первичной структурой белка, т.е. с уникальной последовательностью аминокислот (рис. 23). Уникальность достигается благодаря тому, что тРНК не просто транспортируют нужные аминокислоты к месту синтеза белка, а выстраивают их в строго определенном порядке, который задается порядком расположения триплетов (кодонов) на мРНК. Они (тРНК) фактически узнают, в какое место подставить данную аминокислоту (узнавание происходит благодаря антикодону, который либо находит, либо не находит «свой», комплементарный ему кодон на участке мРНК, захваченной рибосомой в данный момент времени). Обычно сразу насколько рибосом садится на одну молекулу мРНК, одевая еѐ «ѐлочкой» из новосинтезирующихся белковых «веточек». Такая цепочка, состоящая из одной нити мРНК с нанизанными на нее множественными рибосомами, называется полисомой (рис. 24). Кроме триплетов, кодирующих аминокислоты, имеются 3 триплета, UAA, UAG и UGA, которые являются стоп-кодонами на мРНК. Дойдя до них, рибосома прекращает синтез белка; таким образом, стоп-кодон на пути рибосомы обозначает конец белковой цепи (рис. 24). Участок рибосомы, распознающий целые триплеты нуклеотидов РНК, называется рамкой считывания. Ясно, что процесс синтеза белковой молекулы должен начаться точно с первого нуклеотида; если рамка считывания сместится хотя бы на один нуклеотид, все триплеты автоматически изменяют свое содержание. Следует понимать, что в отличие от этой ситуации смещение на один целый триплет ведѐт лишь к потере первой аминокислоты в белке, но не затрагивает другие аминокислоты. 5 Дать краткий ответ на вопрос: что выражает генетический код? Почему код триплетный? Какие молекулы выступают в роли декодирующего механизма? Принцип кодирования определенных аминокислот с помощью соответствующих триплетов нуклеотидов называется генетическим кодом. Триплетность генетического кода вытекает из необходимости закодировать 20 аминокислот с помощью всего лишь 4 нуклеотидных оснований – аденина, гуанина, цитозина и тимина. Если бы каждая аминокислота соответствовала одному нуклеотиду, это позволило бы зашифровать всего 4 аминокислоты. Создание комбинаций из четырех нуклеотидов, взятых по два, дает возможность закодировать 16 аминокислот (42 = 16). Этого также недостаточно для того, чтобы зашифровать все 20 аминокислот. Поэтому в качестве кодирующей единицы берется комбинация из трех нуклеотидов – триплет. Число таких комбинаций равно 64 (43 = 64). Генетический код расшифровывается и реализуется в строении белка с помощью трех видов РНК: матричной, или информационной (мРНК, или иРНК), транспортной (тРНК) и рибосомной (рРНК). 6 Дать краткое определение и формулу центральной догмы молекулярной биологии. Каковы функции ДНК в клетке? Какие синтезы и почему называются матричными? ЦДМБ - Обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Информация о последовательности аминокислот в белках (наследственная, или генетическая информация) хранится и передается в клетке с помощью макромолекул другого типа – нуклеиновых кислот ДНК и РНК. ДНК ответственна за хранение и наследование генетической информации, РНК – за ее реализацию. В результате скоординированной работы трех видов РНК – мРНК, тРНК и рРНК, а также многочисленных внутриклеточных ферментов, осуществляется синтез белковых молекул. ДНК → РНК → белок. Стрелками в данном случае показана передача генетической информации. Как видно из схемы, этот процесс носит однонаправленный характер, исключающий прочтение генетического кода в обратном направлении, т.е. с белка на ДНК. Функции ДНК – хранение и передача наследственной информации. Хранение наследственной информации. Порядок расположения нуклеотидных остатков в молекуле ДНК определяет последовательность аминокислот в молекуле белка. В молекуле ДНК зашифрована вся информация о признаках и свойствах нашего организма. Передача наследственной информации следующему поколению. Эта функция осуществляется, благодаря способности молекулы ДНК к самоудвоению – репликации. Из свободных нуклеотидов бок о бок с цепью ДНК по принципу комплементарности формируется одноцепочечная молекула матричной, или информационной, РНК (мРНК, или иРНК), которая является полной копией одной из цепей ДНК, за исключением того, что вместо тимина в состав РНК по тем же позициям включается урацил (рис. 20). В этом случае транскрибируемый участок молекулы ДНК выступает в роли некоего образца, матрицы для синтеза другой макромолекулы – РНК, а в дальнейшем и белка. Такие синтезы принято называть матричными. 7 Исходя из формулы центральной догмы молекулярной биологии, объяснить, что является молекулярной основой генотипа и фенотипа. Совокупность всех генов данного организма формирует его генотип, а совокупность всех признаков, реализующихся при данном генотипе (т.е., другими словами, совокупность всех белков, синтезируемых в этом организме), объединяют понятием фенотип. 8 Дать определение понятия "репликация", объяснить молекулярный механизм и назначение репликации ДНК.  Репликация, или удвоение, ДНК происходит каждый раз перед делением клетки (за редким исключением). Это необходимое условие для сохранения наследственной информации во всех поколениях клеток и организмов. Наиболее просто организован процесс репликации у прокариот. Единственная кольцевая хромосома бактерий (нуклеоид) реплицируется как одна структурная единица, при этом удвоение начинается в одной стартовой точке репликации (точка origin) и распространяется в обе стороны без перерывов – образуются две вилки репликации, «бегущие» по кольцу в разные стороны. Заканчивается репликация в одной точке терминации (конечная точка репликации), где в итоге сходятся две репликативные вилки. Таким образом, вся ДНК бактерии представляет собой одну единицу репликации – репликон (рис. 40). У эукариотических клеток ДНК имеет полирепликонную организацию, т.е. содержит множество независимых участков репликации, или репликонов (рис. 41). Это существенно ускоряет процесс репликации, хотя и не до абсолютного (теоретически возможного) минимума, т. к. репликация, как правило, начинается не во всех репликонах одновременно. Первым реплицируется эухроматин, последним – конститутивный гетерохроматин центромерных районов хромосом. Существует строгая последовательность репродукции хромосом относительно других хромосом в наборе. Многочисленные ферменты, осуществляющие удвоение ДНК, объединяются в единый репликационный комплекс, под действием которого происходит локальное раскручивание спирали ДНК на две цепи (формирование вилки репликации) и выстраивание свободных нуклеотидов вдоль каждой из цепей по принципу комплементарности. Основным ферментом репликации, как уже говорилось, выступает ДНК-полимераза: она осуществляет подбор комплементарных нуклеотидов. Кроме ДНК-полимеразы здесь работают такие ферменты, как ДНК-праймаза (инициирует репликацию ДНК), ДНК-топоизомераза (препятствует локальной суперспирализации ДНК, обеспечивая возможность посадки ДНК-полимеразы), ДНК-хеликаза (разрывает водородные связи между комплементарными нуклеотидами, раскручивая двойную спираль), ДНК-лигаза (сшивает отдельные нуклеотидные последовательности, полученные в результате матричного синтеза, в одну цепочку) и другие. В результате скоординированной работы ферментов образуются две дочерние молекулы ДНК, полностью идентичные друг другу и материнской молекуле: если в одной цепи мы имеем некую последовательность из 4-х нуклеотидов, то во второй цепи последовательность нуклеотидов будет однозначно детерминирована, так что каждому А в старой цепи будет соответствовать Т в новой цепи, каждому Т старой цепи – А в новой цепи, каждому Г старой цепи – Ц в новой цепи и каждому Ц старой цепи – Г в новой цепи (рис. 42). Такой способ удвоения заложен в самой идее существования ДНК в виде двойной спирали, модель которой была предложена Уотсоном и Криком в 1953 году. При репликации ДНК выполняется также принцип полуконсервативности. Это означает, что каждая из двух цепей ДНК используется в качестве матрицы для образования новой комплементарной цепи. В результате дочерняя ДНК содержит одну новую цепь и одну старую, материнскую. Структура и функции клеточного ядра Основные вопросы: 1 Общеморфологическая характеристика ядерного аппарата эукариотных и прокариотных клеток. Основной генетический материал эукариот (за исключением ДНК митохондрий и пластид) сосредоточен в специальном компартменте – клеточном ядре (рис. 27). Ядро – это достаточно сложно организованная структура, в составе которой можно выделить следующие компоненты: 1) хроматин (хромосомы); 2) ядрышко; 3) ядерная оболочка; 4) ядерный сок (нуклеоплазма, или кариоплазма); 5) ядерный матрикс (белковый скелет ядра). 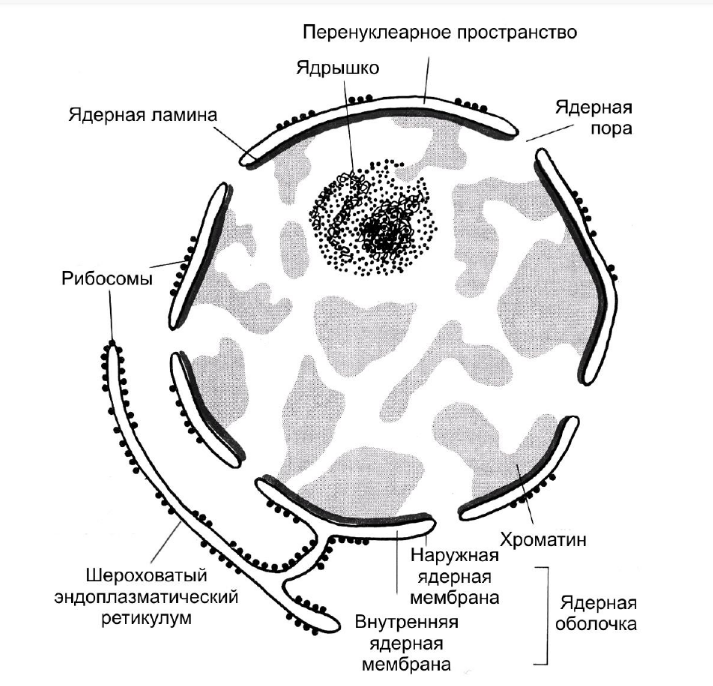 2 Сущность концепции непрерывности хромосом в жизненном цикле клетки. 3 Химический состав хроматина. Что такое ДНП? В химическом смысле хроматин представляет собой комплекс ДНК с ядерными белками, или дезоксирибонуклеопротеидный (ДНП) комплекс. В среднем около 40% массы хроматина приходится на ДНК и около 60% – на белки, хотя это соотношение может изменяться в зависимости от активности клеток. 4 Уровни структурной организации хроматина. Эу- и гетерохроматин. Какие уровни организации хроматина характерны для интерфазного ядра? хроматин в интерфазном ядре (третий уровень компактизации) Первый уровень компактизации хроматина называется нуклеосомной фибриллой, или «бусины на нитке». Основная роль в упаковке ДНК на нуклеосомном уровне принадлежит глобулярным щелочным белкам гистонам (именно благодаря щелочной реакции гистоны связывают и блокируют кислый субстрат, который представляет собой ДНК). Выделяют несколько фракций гистонов: Н1, Н2А, Н2В, Н3 и Н4. При формировании нуклеосомной фибриллы гистоны Н2А, Н2В, Н3 и Н4, взятые каждый по паре, образуют комплексы по 8 штук – октамеры. Вокруг каждого такого октамера нить ДНК совершает 1,75 оборота. Октамер с намотанным на него фрагментом ДНК образует комплекс в виде бусины – нуклеосому. Нуклеосомы связаны между собой свободными участками ДНК – линкерами (от англ. link – связывать). Таким образом, образуется фибрилла толщиной 10 нм, длина которой по сравнению с протяженностью исходной «голой» молекулы ДНК уменьшается в 7 раз. Второй уровень компактизации хроматина получил название нуклеомерной фибриллы. Ещѐ один гистоновый белок – Н1, не задействованный в создании первого уровня компактизации, связывается с линкерными участками ДНК и сближает нуклеосомы между собой, после чего вся нуклеосомная фибрилла еще раз спирализуется: витки спирали также удерживаются гистоном Н1. Получается фибрилла толщиной 30 нм, устроенная либо по типу соленоида (рис. 30, а), либо по типу более крупных «бусин» (супербусин, или нуклеомеров) на более толстой «нитке» (рис. 30, б). Соответственно, выделяют два типа укладки 30- нанометровой фибриллы – соленоидный и нуклеомерный. Независимо от типа укладки, общее укорочение нуклеопротеидной фибриллы (по сравнению с исходной нитью ДНК) на втором уровне компактизации хроматина составляет 40—70 крат. Это последний уровень компактизации, осуществляемый с помощью гистонов. Далее в процессе компактизации участвуют так называемые негистоновые белки. Третий уровень компактизации хроматина можно назвать петельно-хромомерной фибриллой, или хромонемой. Нуклеомерная фибрилла собирается в петли (петельные домены), которые образуют розетки – хромомеры. Размер отдельных петель соответствует одному или нескольким генам. Основания петель содержат определенные последовательности нуклеотидов, которые специфически взаимодействуют с негистоновыми белками ядерного матрикса (скелета) – матриксинами. Участки ДНК петель, связывающиеся с ядерным матриксом, получили название MAR (matrix attachment region), или SAR (scaffold attachment region) и часто обозначаются как MAR/SAR-последовательности. В дальнейшем, обычно при подготовке к митозу, хромомеры максимально компактизуются, сближаются и образуют плотный удлинѐнный нуклеопротеидный тяж – хромонему. При этом хромомеры выявляются по всей длине хромонемы в виде утолщений (рис. 31). Укладка нуклеомерной фибриллы в хромонему позволяет достичь 600—700-кратного укорочения фибриллы по сравнению с «голой» ДНК. Четвертый уровень компактизации хроматина – хроматидный, или хромосомный – образуется в период митотического деления клетки. При этом хромонема спирализуется с образованием удлинѐнных оптически плотных телец – хроматид. Следует помнить, что перед делением ДНК удваивается (это происходит задолго до начала формирования четвертого уровня компактизации), а это значит, что каждая интерфазная хромосома после удвоения будет состоять не из одной, а из двух хромонем, которые затем, уже при переходе к митозу, конденсируются до уровня хроматид. Хромосома, состоящая из двух максимально компактизованных хроматид, окончательно формируется к моменту метафазы (вторая стадия митоза, см. ниже) и поэтому называется метафазной хромосомой (рис. 32). В интерфазном ядре можно выделить две фракции хроматина, различающиеся в структурном и функциональном отношениях. Рыхлое состояние хроматина в виде раскрученных нитей ДНП, максимально освобожденных от компактизующих белков, является единственно возможным для осуществления процессов транскрипции и репликации. Такой активированный хроматин, доступный для считывания информации, получил определение эухроматина. В состоянии эухроматина в ядре содержатся именно те гены, активность которых необходима данному типу клеток в данный момент. Весь остальной объем генетического материала (а его большинство) пребывает в клетке в неактивном состоянии, не работает и заблокирован с помощью компактизующих белков. Хроматин, находящийся в таком конденсированном нерабочем состоянии, называется гетерохроматином. Выделяют две разновидности гетерохроматина – факультативный («необязательный») и конститутивный («обязательный»). В состоянии факультативного гетерохроматина могут находиться смысловые последовательности, т.е. гены, которые «молчат» временно, например, за ненадобностью в данный момент или в данном типе клеток. Такие «молчащие» гены потенциально могут деконденсироваться и начать транскрибироваться. Конститутивный же гетерохроматин – это та существенная доля ДНК, которая в принципе неактивна в генетическом отношении (не кодируют белки) и не переводится в состояние эухроматина ни при каких обстоятельствах. Речь идет о той самой сателлитной ДНК, которая составляет большую часть «бессмысленной», избыточной ДНК генома эукариот,о чем уже упоминалось выше. Напомним, что сателлитной ДНК приписывают ,в частности, структурную функцию – считается, что она способствует закреплению хромосом на скелетных структурах ядра. 5 Какие проявления транскрипции мРНК можно видеть в световой и электронный микроскоп? 6 Строение хромосом типа ламповых щеток и политенных хромосом, соответствие их деталей хроматиновым структурам обычных ядер. 7 Строение и функции ядрышка. Объяснить сущность процессинга рРНК. Совокупность рибосомных генов и продуктов их активации, как промежуточных, так и конечных, включая готовые рибосомные субъединицы, и составляет зону повышенной плотности – ядрышко. В этой части ядра происходит транскрипция генов, кодирующих рибосомную РНК (рибосомных генов, или р-генов), созревание, или процессинг, рРНК и сборка рибосомных субъединиц из фрагментов рРНК и рибосомных белков. Гены рибосомной ДНК группируются в кластеры, называемые ядрышковыми организаторами (ЯО), или ядрышкообразующими районами (ЯОР), которые располагаются на определенных участках одной или нескольких ядрышкообразующих хромосом. Морфологически в ядрышке различают три основные зоны: фибриллярный центр (или несколько центров), окруженный плотным фибриллярным и гранулярным компонентами (рис. 37). Установлено, что в фибриллярном центре (ФЦ) ядрышка локализованы сами рибосомные гены, а также белки, необходимые для их транскрипции и созревания. Считается, что транскрипция генов рРНК происходит на периферии фибриллярного центра, то есть на границе его с плотным фибриллярным компонентом (ПФК). Таким образом, сам плотный фибриллярный компонент представлен массой растущих и созревающих цепей рРНК и ассоциированными с ними белками. Гранулярный компонент (ГК) образован большим количеством готовых рибосомных субчастиц. Кроме того, в составе ядрышка, как правило, имеются глыбки конститутивного гетерохроматина (так называемый околоядрышковый гетерохроматин), с помощью которого ядрышко прикрепляется к ядерной оболочке или к элементам ядерного скелета (рис. 37). Помимо чисто структурной функции, некоторые исследователи отводят околоядрышковому гетерохроматину и регуляторную роль. Благодаря безнуклеосомной организации ядрышкового хроматина возможна быстрая мобилизация «молчащих» р-генов в пределах каждого отдельного ядрышка и вовлечение их в синтез рибосом. За счет этого размер и плотность ядрышка может постоянно меняться. Однако не вся нуклеотидная последовательность ДНК, входящая в состав гена, содержит полезную информацию для синтеза РНК. Кодирующие последовательности внутри гена называются экзонами, а разделяющие их некодирующие последовательности – интронами. Молекула РНК, только что синтезированная с гена, носит название первичного транскрипта, или пре-РНК. Для того чтобы первичный транскрипт превратился в зрелую РНК, он должен подвергнуться процессингу – серии посттранскрипционных изменений. Важнейшая часть процессинга – сплайсинг – вырезание некодирующих последовательностей (интронов) и сращивание кодирующих (экзонов) (рис. 46). Такая прерывистая, экзон- интронная организация гена – эволюционное приобретение эукариотических клеток. 8 Строение эукариотической рибосомы: субъединицы, параметры молекул РНК, белки. 9 Что такое амплификация ядрышковой ДНК? Где известна и для чего она нужна? 10 Ядерный матрикс и ядерная оболочка: их строение и значение в организации работы хроматина.  Для поддержания пространственной структуры ядра существует особая система, называемая ядерным скелетом, или ядерным матриксом. Ядерный матрикс включает в себя ламину, подстилающую ядерную мембрану изнутри, и систему белковых нитей, пронизывающих ядро и формирующих в его пространстве разветвленную сеть. Как правило, такие нити одним концом крепятся к ламине (с помощью других специальных белков), а другим концом связываются с определенными участками хромосом. Таким образом, матрикс создает некий каркас, объединяющий и структурирующий в пространстве все ядерные компоненты: хроматин, ядрышко, ядерную оболочку. Оболочка ядра образована двумя мембранами. Выделяют внутреннюю ядерную мембрану, обращенную к полости ядра, и наружную ядерную мембрану, обращенную к цитоплазме. Наружная мембрана переходит непосредственно в мембрану эндоплазматической сети, или эндоплазматического ретикулума (ЭПС, или ЭПР, или ЭР). Между внутренней и наружной мембранами образуется узкое пространство, названное перинуклеарным (рис. 38, А), которое, соответственно, сообщается с полостью каналов и цистерн эндоплазматического ретикулума. С внутренней стороны ядерная оболочка (ее внутренняя мембрана) подстилается плотной пластиной – ламиной, которая поддерживает ядерную мембрану и контактирует с хромосомами. Ламина является компонентом ядерного скелета, или ядерного матрикса, она образована специальными белками ламинами. У высших эукариот в состав плотной пластинки входят ламины трех типов – А, В и С, которые весьма незначительно отличаются друг от друга по аминокислотной последовательности, то есть характеризуются высоким процентом гомологии. Степень развития плотной пластины может варьировать в широких пределах: от едва заметной (20—80 нм) субмембранной сети до мощного (250—300 нм) сотового слоя ядра у некоторых простейших. 11 Строение и функции ядерных пор.  Ядерная оболочка имеет многочисленные поровые комплексы для сообщения с цитоплазмой и транспорта веществ, в том числе для переноса из ядра в цитоплазму молекул мРНК в связке со специальными белками. Ядерная пора представляет собой комплекс из 17 белков, где центральная белковая глобула играет роль канала, а остальные 16 образуют два кольца по 8 глобул в каждом, прикрепленные к наружной и внутренней ядерной мембране соответственно и фиксирующие, таким образом, центральную глобулу в центре поры (рис. 38). Все макромолекулы, проходящие через ядерные поры, имеют в своѐм составе особую полипептидную (аминокислотную) последовательность, получившую название сигнал ядерного импорта и предназначенную для распознавания порой транспортируемого вещества. После преодоления ядерной поры эта сигнальная последовательность отсоединяется от макромолекулы. Дополнительные вопросы: 12 Что такое ген? Это понятие молекулярное, структурное, функциональное или генетическое? 13 Почему хроматин отличается высокой базофилией? 14 Раскрыть представление о прерывистой структуре гена: экзон-интронная организация гена, особенности процессинга мРНК, механизм сплайсинга. 15 В нервных клетках ядра обычно крупные и бледные (ДНК выявляется с трудом), в эритроцитах птиц, рыб, напротив - ядра мелкие и очень плотные. Что можно сказать о структуре хроматина и функции этих ядер? 16 Генетическая конституция человека несравненно сложнее, чем у лягушки или рыбы. В то же время у некоторых амфибий и древних рыб нормальное количество ядерной ДНК в 10-50 раз больше, чем у человека и других млекопитающих (2с человека - 6пг - пикограмм ДНК, лягушки - 11пг, тритона - 74пг, амфиумы - 108пг, саламандры - 340пг). Почему? 17 Каков путь переноса субъединиц рибосом из ядрышка в цитоплазму? 18 Какова роль рРНК в организации или функционировании рибосомы? 19 Почему гены рРНК в ядрышковом организаторе имеют многократную повторность? 20 Как с помощью авторадиографии выявить место синтеза, направление и скорость перемещения синтезированной РНК? 21 Какими цитохимическими методами можно выявить ДНК и измерить ее количество в клеточном ядре? |