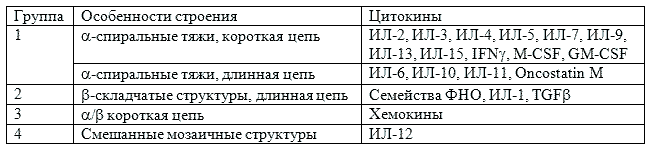
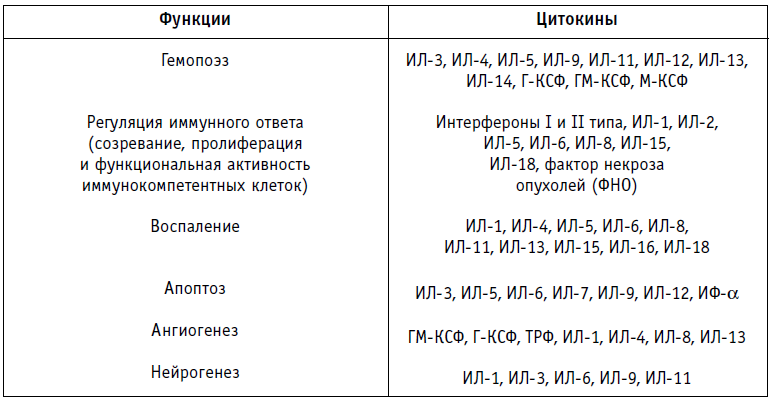
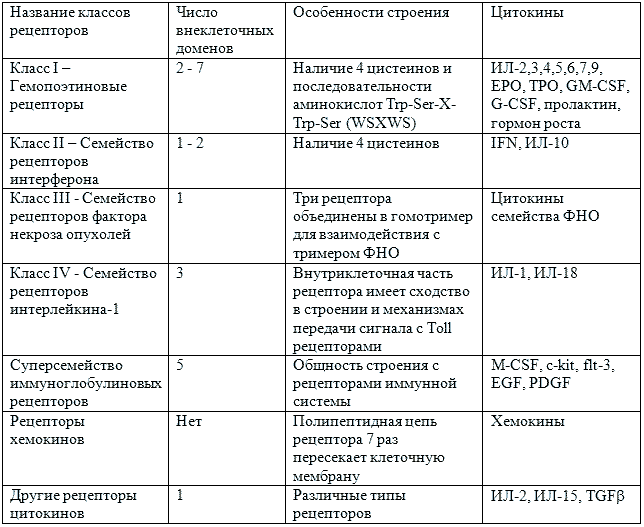
Цитокины
 Скачать 1.35 Mb. Скачать 1.35 Mb.
|

Общая характеристика цитокинов Механизм действия цитокинов Механизм нарушения Интерлейкины Интерфероны TNF: Фактор некроза опухолей Колониестимулирующие факторы
Общие свойства цитокинов: 1. Цитокины являются полипептидами или белками, часто гликозилированными, большинство из них имеют ММ от 5 до 50 кДа. Биологически активные молекулы цитокинов могут состоять из одной, двух, трех и более одинаковых или разных субъединиц. 2. Цитокины не имеют антигенной специфичности биологического действия. Они влияют на функциональную активность клеток, принимающих участие в реакциях врожденного и приобретенного иммунитета. Тем не менее, воздействуя на Т- и В-лимфоциты, цитокины способны стимулировать индуцированные антигенами процессы в иммунной системе. 3. Для генов цитокинов существуют три варианта экспрессии: а) стадиоспецифическая экспрессия на определенных стадиях эмбрионального развития, б) конститутивная экспрессия для регуляции ряда нормальных физиологических функций, в) индуцибельный тип экспрессии, характерный для большинства цитокинов. Действительно, большинство цитокинов вне воспалительной реакции и иммунного ответа не синтезируются клетками. Экспрессия генов цитокинов начинается в ответ на проникновение в организм патогенов, антигенное раздражение или повреждение тканей. Одними из наиболее сильных индукторов синтеза провоспалительных цитокинов служат патоген-ассоциированные молекулярные структуры. Для запуска синтеза Т-клеточных цитокинов требуется активация клеток специфическим антигеном с участием Т-клеточного антигенного рецептора. 4. Цитокины синтезируются в ответ на стимуляцию короткий промежуток времени. Синтез прекращается за счет разнообразных механизмов ауторегуляции, включая повышенную нестабильность РНК, и за счет существования отрицательных обратных связей, опосредуемых простагландинами, кортикостероидными гормонами и другими факторами. 5. Один и тот же цитокин может продуцироваться различными по гистогенетическому происхождению типами клеток организма в разных органах. 6. Цитокины могут быть ассоциированными с мембранами синтезирующих их клеток, обладая в виде мембранной формы полным спектром биологической активности и проявляя свое биологическое действие при межклеточном контакте. 7. Биологические эффекты цитокинов опосредуются через специфические клеточные рецепторные комплексы, связывающие цитокины с очень высокой аффинностью, причем, отдельные цитокины могут использовать общие субъединицы рецепторов. Рецепторы цитокинов могут существовать в растворимой форме, сохраняя способность связывать лиганды. 8. Цитокины обладают плейотропностью биологического действия. Один и тот же цитокин может действовать на многие типы клеток, вызывая различные эффекты в зависимости от вида клеток-мишеней. Плейотропность действия цитокинов обеспечивается экспрессией рецепторов цитокинов на разных по происхождению и функциям типах клеток и проведением сигнала с использованием нескольких разных внутриклеточных мессенджеров и транскрипционных факторов. 9. Для цитокинов характерна взаимозаменяемость биологического действия. Несколько разных цитокинов могут вызывать один и тот же биологический эффект либо обладать похожей активностью. Цитокины индуцируют либо подавляют синтез самих себя, других цитокинов и их рецепторов. 10. В ответ на активационный сигнал происходит синтез клетками одновременно нескольких цитокинов, участвующих в формировании цитокиновой сети. Биологические эффекты в тканях и на уровне организма зависят от присутствия и концентрации других цитокинов с синергичным, аддитивным или противоположным действием. 11. Цитокины могут влиять на пролиферацию, дифференцировку и функциональную активность клеток-мишеней. 12. Цитокины действуют на клетки различными путями: аутокринно – на клетку, синтезирующую и секретирующую данный цитокин; паракринно – на клетки, расположенные вблизи клетки-продуцента, например, в очаге воспаления или в лимфоидном органе; эндокринно – дистантно на клетки любых органов и тканей после попадания в циркуляцию. В последнем случае действие цитокинов напоминает действие гормонов. Один и тот же цитокин может продуцироваться различными по гистогенетическому происхождению типами клеток организма в разных органах и действовать на многие типы клеток, вызывая различные эффекты в зависимости от вида клеток-мишеней. 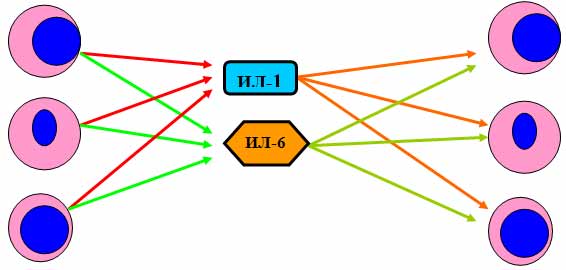 Три варианта проявления биологического действия цитокинов. 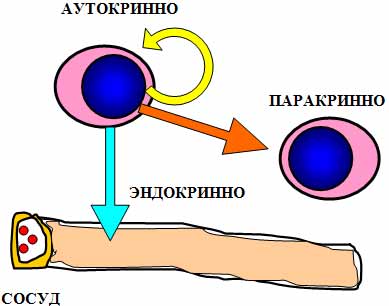 По-видимому формирование системы цитокиновой регуляции эволюционно проходило вместе с развитием многоклеточных организмов и было обусловлено необходимостью образования посредников межклеточного взаимодействия, к которым могут быть причислены гормоны, нейропептиды, молекулы адгезии и некоторые другие. Цитокины в этом плане являются наиболее универсальной системой регуляции, так как способны проявлять биологическую активность как дистантно после секреции клеткой-продуцентом (местно и системно), так и при межклеточном контакте, будучи биологически активными в виде мембранной формы. Этим система цитокинов отличается от молекул адгезии, выполняющих более узкие функции только при непосредственном контакте клеток. В то же время система цитокинов отличается от гормонов, которые в основном синтезируются специализированными органами и оказывают действие после попадания в систему циркуляции. Роль цитокинов в регуляции физиологических функций организма может быть разделена на 4 основных составляющих: 1. Регуляция эмбриогенеза, закладки и развития органов, в т.ч. органов иммунной системы. 2. Регуляция отдельных нормальных физиологических функций. 3. Регуляция защитных реакций организма на местном и системном уровне. 4. Регуляция процессов регенерации тканей. 2.Интерлейкины Это большая группа цитокинов (от ИЛ-1 до Ил-18), синтезируемых в основном T-клетками, но в некоторых случаях также мононуклеарными фагоцитами или другими тканевыми клетками. Интерлейкины обладают разнообразными функциями, но большинство их стимулирует другие клетки для деления или дифференцировки, при этом каждый из них действует на отдельную, ограниченную группу клеток, экспрессирующих специфичные для данного интерлейкина рецепторы. Это растворимые пептиды, сильные иммунорегуляторы локального действия ; активируют Т-клетки. Функции интерлейкинов связаны с активностью других физиологически активных пептидов и гормонов: эндотелина, пролактина, брадикинина. Интерлейкин 1 (англ. Interleukin-1, IL-1) — цитокин, медиатор воспаления и иммунитета, синтезируется многими клетками организма, в первую очередь активированными макрофагами, кератиноцитами, стимулированными B-клетками и фибробластами. Интерлейкин-1 был одним из первых открытых интерлейкинов. Первоначально он был описан как фактор, вызывающий повышение температуры, контролирующий активностьлейкоцитов, увеличивающий количество клеток костного мозга и приводящий к дегенерации суставов. Позже было обнаружено, что существует два сходных интерлейкина 1: альфа и бета. Кроме интерлейкинов 1α и 1β в группу интерлейкинов 1 относят антагонист рецептора интерлейкина 1, а также интерлейкины -18 и -33. Все они обладают определённым сходством структуры. Кроме этого, было обнаружено ещё 6 факторов, отнесённых к этому надсемейству. Интерлейкин-1α и интерлейкин-1β синтезируются в виде предшественников и превращаются в зрелые белки после отщепления пропептида либо протеазой каспаза-1, либо т.н. интерлейкин-1-конвертирующим ферментом (ICE). Оба белка имеют молекулярную массу 18 кДа. Структура обоих интерлейкинов включает 12-14 β-складок, образующих бочкообразный или цилиндрический белок.
Интерлейкин 18 (англ. interleukin 18, IL-18) — провоспалительный цитокин, принадлежащий к семейству интерлейкина 1. Синтезируетсямакрофагами и другими клетками организма. Играет значительную роль в инфекционных и аутоиммунных заболеваниях. Был впервые открыт в1995 году в качестве интерферон-гамма-индуцирующего фактора, который увеличивал активность клеток ЕК в селезёнке, причём его активность выше, чем у интерлейкина 12. Интерлейкин-18 синтезируется в виде белка-предшественника, состоящего из 192 аминокислот, который под действием интерлейкин-1-конвертирующего фермента превращается в зрелый белок из 157 аминокислот. После секреции из клетки-продуцента интерлейкин-18 связывается либо с интерлейкин-18-связывающим белком, который инактивирует его, либо с интерлейкин-18-рецепторным комплексом. Последний включаетрецептор интерлейкина-18 (IL-1R5), входящий в надсемейство интерлейкин-1/толл-подобных рецепторов, и интерлейкин-18-дополняющий белок (IL-1R7). После формирования лиганд-рецепторного комплекса к нему присоединяется адаптерный белок MyD88 и киназа IRAK1, запускающая сигнальный путь, активирующий провоспалительный фактор трансрипции NF-κB. Интерлейкин 18 (интерферон-гамма-индуцирующий фактор)  Интерлейкин 33 (англ. interleukin 33, IL-33) — цитокин, принадлежащий к семейству интерлейкина 1, обладает сходством с интерлейкином 1 ифактором роста фибробластов. Экспрессируется многими клетками организма, его уровень строго коррелирует с уровнем воспаления в ткани. В отличие от провоспалительного интерлейкина-1 интерлейкин-33 обладает иммунорегуляторными свойствами. Интерлейкин-33 синтезируется в виде предшественника с молекулярной массой 30 кДа, после отщепления пропептида под действием ферментакаспаза-1 превращается в зрелый белок массой 18 кДа. Зрелый белок секретируется и регулирует активность Т-хелперов 2 типа. Однако, синтезированный интерлейкин-33 может и не проходить стадию созревания. В этом случае он может напрямую действовать как фактор транскрипции благодаря наличию ядерного сигнала в пропептиде.  Ил-2 вырабатывается Т-лимфоцитами, гл. обр. Thl, а также цитотоксическими лимфоцитами (CD8+) 1 порядка. Он активирует дифференцировку Thl и Т-киллеров, стимулирует NK и синтез иммуноглобулинов В-лимфоцитами. Ил-3 продуцируется Т-лимфоцитами и стволовыми клетками. Является ростовым фактором стволовых и ранних предшественников гемопоэтических клеток. Ил-4 продуцируется гл. обр. Th2. Он стимулирует дифференцировку Th0 в Th2, стимулирует синтез иммуноглобулинов В-лимфоцитами, подавляет генерацию цитотоксических лимфоцитов, NK, а также продукцию ИФН-g и противоопухолевую активность макрофагов. Ил-5 синтезируется Th2. Способстует пролиферации и дифференцировке стимулированных В-лимфоцитов, усиливает продукцию IgA, активирует эозинофилы. Ил-6 вырабатывается макрофагами, Т- и В-лимфоцитами. Стимулирует пролиферацию тимоцитов, В-лимфоцитов, активирует предшественников цитотоксических лимфоцитов, гранулоцитов и макрофагов, стимулирует образование гепатоцитами белков острой фазы, оказывает провоспалительное действие, обеспечивает взаимосвязь иммунной, нервной и эндокринной систем. ИЛ-7 продуцируется стромальными клетками костного мозга. Является ростовым фактором пре-В- и пре-Т-лимфоцитов. ИЛ-8 синтезируется моноцитами, макрофагами, фибробластами. Вызывает миграцию нейтрофилов и базофилов в очаг воспаления и их дегрануляцию, выделение супероксидного радикала. Стимулирует ангиогенез. ИЛ-9 продуцируется гл. обр. Т-лимфоцитами. Стимулирует пролиферацию Т-лимфоцитов, активирует тучные клетки, усиливает эффекты эритропоэтина. ИЛ-10 синтезируется Th2 а также цитотоксическими Т-лимфоцитами второго порядка и макрофагами. Стимулирует пролиферацию и дифференцировку В-лимфоцитов, подавляет синтез ИЛ-2 и ИФН-у клетками Thl, угнетает продукцию провоспалительных цитокинов. ИЛ-11 продуцируется стромальными клетками костного мозга. Стимулирует деление и дифференцировку предшественников гемопоэза, колониеобразование мегакариоцитов, увеличивает количество тромбоцитов и эритроцитов в периферической крови. Угнетает продукцию провоспалительных цитокинов. ИЛ-12 продуцируют моноциты, макрофаги и, в меньшей степени, В-лимфоциты и дендритные клетки. Стимулирует рост и дифференцировку Th (Th0 => Thl), Т-киллеров, NK. Индуцирует продукцию ИФН-g Т-лимфоцитами и NK, угнетает апоптоз Thl, синтез IgE. Вместе с ИЛ-4 регулирует баланс Thl и Th2. ИЛ-13 синтезируется Th2. Стимулирует рост и дифференцировку В-лимфоцитов, подавляет функцию моноцитов/макрофагов, в частности секрецию провоспалительных цитокинов. ИЛ-14 продуцируется в основном Т-лимфоцитами. Усиливает пролиферацию В-лимфоцитов и подавляет продукцию иммуноглобулинов. ИЛ-15 вырабатывается моноцитами, эпителиоцитами и гладкомышечными клетками. По действию на Т-лимфоциты ИЛ-15 сходен с ИЛ-2, что объясняется способностью специфически связываться с ИЛ-2-рецепторами. Активирует NK и В-лимфоциты. ИЛ-16 синтезируется эозинофилами и CD8+ Т-лимфоцитами. Является хемоаттрактантом для CD4+ лимфоцитов. ИЛ-17 продуцируется активированными CD4 Т-лимфоцитами. Основными клетками-мишенями цитокина являются эпителиоциты, эндотелиоциты и фибробласты. Он усиливает выработку ИЛ-б, ИЛ-8, гранулоцитарного КСФ, простатландина Е2, увеличивает экспрессию ICAM-1, стимулирует активность фибробластов. 3.Интерфероны Эти цитокины особенно важны в сдерживании некоторых вирусных инфекций, они образуются на ранней стадии инфекции и создают первую линию защиты против большинства вирусов. Противовирусный препарат интерферон обладает лечебной и профилактической активность при вирусных инфекциях. Генноинженерный интерферон - интерлок- используют для лечения вирусных заболеваний глаз, генноинженерный интерферон -реаферон- при вирусном гепатите. Интерфероны (IFN) - это ряд антивирусных агентов широкого спектра действия. Существует по меньшей мере 14 альфа-интерферонов, которые продуцируются лимфоцитами, тогда как фибробласты, а возможно, и другие клетки продуцируют бета-интерферон. Образование гамма-интерферона индуцируется вирусами. При вирусной инфекции клетки синтезируют интерферон и секретируют его в межклеточное пространство, где он связывается со специфическими рецепторами соседних незараженных клеток. Связанный интерферон оказывает противовирусное действие. Конечный результат состоит из образования барьера из устойчивых к вирусу неинфицированных клеток вокруг очага инфекции, чтобы ограничить ее распространение. Однако следует подчеркнуть, что интерфероны играют большую роль именно в борьбе с вирусами, а не в предотвращении вирусных инфекций. ИнтерфероныИФ-альфаиИФ-бетасинтезируют клетки, инфицированные вирусами. ИнтерферонИФ-гаммавыделяют некоторые активированныеT-лимфоцитыиНК-клетки. Его образование не индуцируется вирусами. ИФ-гамма оказывает многочисленные иммунорегуляторные эффекты. Противовирусное и антипролиферативное действие гамма-интерферона менее выражено, чем у ИФ-альфа и ИФ-бета. Человеческий интерферон-α  Человеческий интерферон-β  Человеческий интерферон-γ  При заражении клетки вирус начинает размножаться. Клетка-хозяин одновременно с этим начинает продукцию интерферона, который выходит из клетки и вступает в контакт с соседними клетками. Хотя интерферон не обладает прямым противовирусным действием, он способен вызывать такие изменения в клетках, которые препятствуют размножению вируса, формированию вирусных частиц и дальнейшему его распространению. Интерферон действует в нескольких направлениях. Во-первых, он оказывает влияние на клетки, соседние с инфицированной, запуская в них цепь событий, приводящих к подавлению синтеза вирусных белков и в некоторых случаях сборки и выхода вирусных частиц (путём активации олигоаденилатциклазы). В ответ на воздействие интерферона клетки вырабатывают большое количество протеинкиназы R. Этот фермент фосфорилирует фактор инициации трансляции eIF-2, фосфорилированный eIF-2 формирует неактивный комплекс с другим фактором, eIF-2B. В результате уровень белкового синтеза в клетке снижается. После протеинкиназы R активируется синтез рибонуклеазы L, которая расщепляет клеточные РНК и ещё больше снижает уровень белкового синтеза. В целом, интерферон-зависимое подавление трансляции является губительным как для вируса, так и для клетки-хозяина. Помимо влияния на трансляцию, интерфероны способны активировать сотни других генов (они известны как гены, стимулируемые интерфероном), играющих роль в защите клетки от вирусов[2][3]. Кроме того, интерферон лимитирует распространение вирусных частиц путём активации белка p53, что ведёт к апоптотической смерти инфицированной клетки[4][5]. Вторым направлением действия интерферонов является стимуляция иммунной системы для борьбы с вирусами. Интерферон повышает синтез молекул главного комплекса гистосовместимости I и II классов и активирует иммунопротеасому. Высокий уровень молекул главного комплекса гистосовместимости I класса обеспечивает эффективную презентацию вирусных пептидовцитотоксическим Т-лимфоцитам и натуральным киллерам, а иммунопротеасома осуществляет процессинг вирусных пептидов, предшествующий презентации. Высокий уровень молекул главного комплекса гистосовместимости II класса обеспечивает презентацию вирусных антигенов Т-хелперам. Т-хелперы, в свою очередь, выделяют цитокины, которые координируют активность других клеток иммунной системы. Некоторые виды интерферонов, например интерферон-γ, могут прямо стимулировать клетки иммунной системы, такие как макрофаги и натуральные киллеры. Образование интерферона могут стимулировать не только интактные вирусы, но и различные другие агенты, например некоторые инактивированные вирусы, двухцепочечные РНК, синтетические двухцепочечные олигонуклеотиды и бактериальные эндотоксины. Биологическая активность интерферона очень высока. У мышиного интерферона она составляет 2·109 ед./мг., а одна единица снижает образование вирусов примерно на 50 %. Это означает, что достаточно одной молекулы интерферона, чтобы сделать клетку резистентной к вирусной инфекции. Показано, что молекулы интерферона должны оказывать действие на клетку в течение минимум четырех часов, для того, чтобы в клетке начались процессы борьбы с вирусом, таким образом, многие специалисты не считают эффективным интраназальное применение интерферона для профилактики ОРВИ[6]. Тем не менее, последние исследования показывают, что интерферон, примененный на слизистую оболочку, может действовать в качестве иммунологического адъювантапротив вируса гриппа, усиливая специфический ответ иммунной системы.[7]. В США проводятся клинические испытания вакцины против гриппа, которая использует интерферон в качестве адъюванта[8]. Интерферон вызывает и целый ряд других биологических эффектов, в том числе подавляет размножение клеток. Недавние исследования показали, что в определённых условиях он может препятствовать развитию злокачественных новообразований. Установлено также, что интерферон действует на иммунную систему и вызывает изменение клеточных мембран. Таким образом, интерфероновая система, вероятно, может играть важную роль в защите организма отвирусов. 4.TNF, ФНО (Фактор некроза опухолей) ФНО-a и ФНО-b - два близких белка (гомологичны около 30% аминокислотных остатков) - проявляют сходную активность по отношению к воспалительной реакции, иммунным и опухолевым процессам. ФНО-а, впервые обнаруженный в сыворотке мышей, которым вводили бактериальные продукты, индуцирует некроз опухолевых клеток. ФНО-b, или лимфотоксин, был найден в лимфатических узлах иммунизированных крыс.Источником ФНО-а является активированный макрофаг,ФНО-b - активированная T-клетка. Оба фактора через одинаковые специфические рецепторы TNF клеточной поверхности вызывают лизис клеток лимфомы, некроз саркомы, индуцированной метилхолантреном, активируют полиморфноядерные лейкоциты, проявляют антивирусную активность. ФНО-альфа (называемый также кахектином) - этопироген. Играет важную роль в патогенезесептического шока, вызванного грамотрицательными бактериями. Под влиянием ФНО-альфа резко увеличивается образование макрофагами и нейтрофиламиперекиси водородаи другихсвободных радикалов. При хроническом воспалении ФНО-альфа активирует катаболические процессы и тем самым способствует развитиюкахексии- симптома многих хронических заболеваний. При изучении различных продуктов, секретируемых активированнымимакрофагами, был получен фактор, который лизировал большой наборопухолевых клетокin vivo и in vitro. По основному биологическому эффекту он получил название - фактор некроза опухолей. В параллельных исследованиях из культур активированныхТ-клетоквыделили другой фактор, также обладавший литической активностью по отношению к чужеродным клеткам. По типу клеток, продуцирующих этот фактор, его стали обозначать каклимфотоксин. Детальное изучение этих факторов вскрыло близкое структурное и функциональное сходство между ними. Настоящее их наименование -фактор некроза опухолей-альфа (ФНО-альфа)ифактор некроза опухолей-бета (ФНО-бета, лимфотоксин). 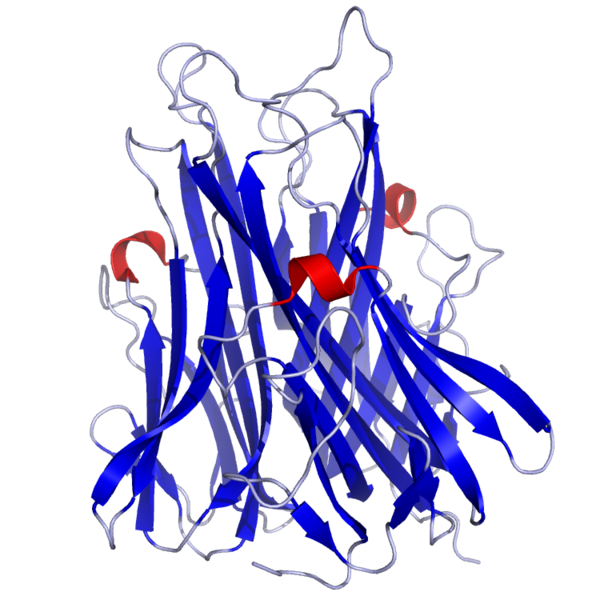 5. Колониестимулирующие факторы Колониестимулирующие факторы - гормоны, стимулирующие образованиемоноцитови нейтрофиловвкостном мозге. При исследовании культуры кроветворных клеток показано, что для размножения и дифференцирования клеток необходимы специфическиефакторы роста. Факторы, поддерживающиегемопоэзв такой культуре, этогликопротеиныи обычно их называют колониестимулирующими факторами, или КСФ. Из все возрастающего числа КСФ, которые были идентифицированы, одни циркулируют в крови и действуют какгормоны, в то время как другие играют роль локальныххимических медиаторов. Этицитокины(колониестимулирующие факторы) участвуют в регуляции деления и дифференцировки стволовых клеток костного мозга и клеток - предшественников лейкоцитов крови. Балансом различных КСФ в определенной мере обусловлено соотношение между различными типами образующихся в костном мозге лейкоцитов. Некоторые КСФ стимулируют дальнейшую дифференцировку клеток и вне костного мозга. Из КСФ гормонального типа лучше всего изученэритропоэтин, который вырабатывается в почкахи регулируетэритропоэз(образованиеэритроцитов). За выживание и пролиферацию плюрипотентных стволовых клеток и большинства типов их коммитированных потомков эритроидного ряда ответственен второй колониестимулирующий фактор -интерлейкин 3(ИЛ-3).Было выявлено также четыре различных КСФ, стимулирующих в культуре клеток формирование колонийнейтрофиловимакрофагов. Эти КСФ синтезируются клетками разного типа, в том числеэндотелиальными клетками,фибробластами,макрофагамиилимфоцитами. Это выше упомянутый интерлейкин 3 и более избирательныеГМ-КСФ(длягранулоцитовимакрофагов),Г-КСФ(для гранулоцитов) иМ-КСФ(для макрофагов). Как и эритропоэтин, все эти КСФ являются гликопротеинами. Их воздействие на клетки-предшественники заключается не только в запуске механизма образования дифференцированных колоний, но и в активации специализированных функций (таких, какфагоцитози убивание клеток-мишеней) у клеток с законченной дифференцировкой. Механизмы действия цитокинов Различают интракринный, аутокринный, паракринный и эндокринный механизмы действия цитокинов. 1. Интракринный механизм - действие цитокинов внутри клетки-продуцента; связывание цитокинов со специфическими внутриклеточными рецепторами. 2. Аутокринный механизм - действие секретируемого цитокина на саму секретирующую клетку. Например, интерлейкины-1, -6 -18, ФНОα являются аутокринными активирующими факторами для моноцитов/макрофагов. 3. Паракринный механизм - действие цитокинов на близкорасположенные клетки и ткани. Например, ИЛ-1, -6 -12 и -18, ФНОα, продуцируемые макрофагом, активируют Т-хелпер (Th0), распознающий антиген и МНС макрофага (Схема аутокринно-паракринной регуляции иммунного ответа). 4. Эндокринный механизм - действие цитокинов на расстоянии от клеток-продуцентов. Например, ИЛ-1, -6 и ФНОα, помимо ауто- и паракринных воздеиствий могут оказывать дистантное иммунорегуляторное действие, пирогенный эффект индукцию выработки белков острой фазы гепатоцитами, симптомы интоксикации и мультиорганные поражения при токсико-септических состояниях. Аутокринно-паракринная регуляция иммунного ответа 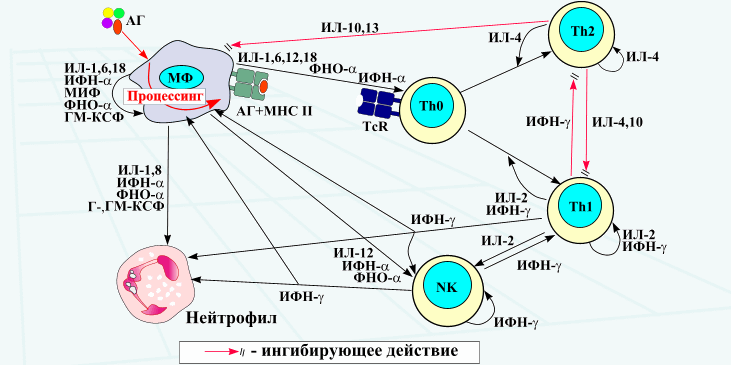 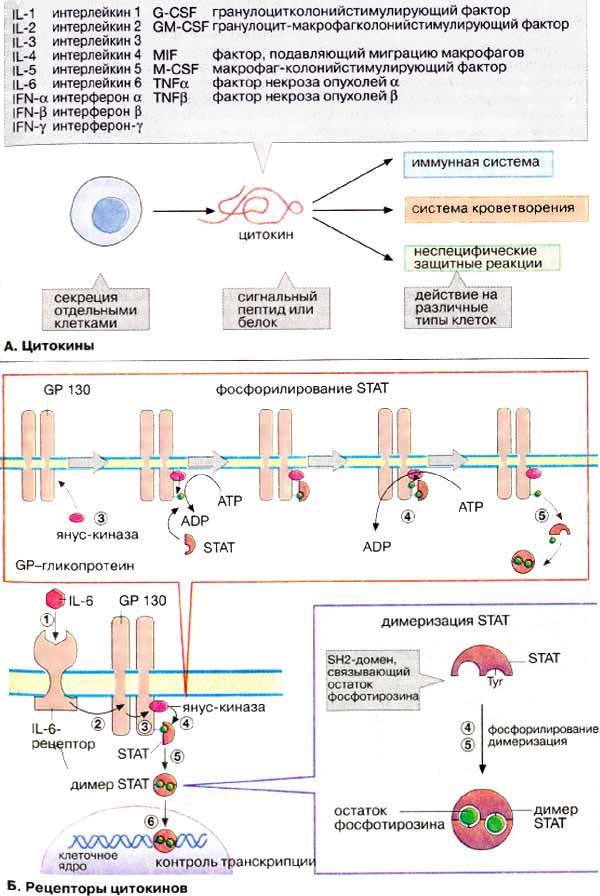 Механизм нарушения Многие тяжелые заболевания приводят к значительному повышению уровня ИЛ-1 и ФНО-альфа. Эти цитокины способствуют активации фагоцитов, их миграции в место воспаления, а также высвобождению медиаторов воспаления - производных липидов, то есть простагландина Е2, тромбоксанов и фактора активации тромбоцитов. Кроме того, они прямо или опосредованно вызывают расширение артериол, синтез адгезивных гликопротеидов, активируют Т- и В-лимфоциты. ИЛ-1 запускает синтез ИЛ-8, способствующего хемотаксису моноцитов и нейтрофилов и выходу ферментов из нейтрофилов. В печени снижается синтез альбумина и усиливается синтез белков острой фазы воспаления, включая ингибиторы протеаз, компоненты комплемента, фибриноген, церулоплазмин, ферритин и гаптоглобин. Уровень С-реактивного белка, который связывается с поврежденными и погибшими клетками, а также некоторыми микроорганизмами, может повышаться в 1000 раз. Возможно также значительное повышение концентрации амилоида A в сыворотке и его отложение в различных органах, приводящее к вторичному амилоидозу. Важнейшим медиатором острой фазы воспаления является ИЛ-6, хотя ИЛ-1 и ФНО альфа тоже могут вызывать описанные изменения функции печени. ИЛ-1 и ФНО альфа усиливают влияние друг друга на местные и общие проявления воспаления, поэтому сочетание этих двух цитокинов даже в небольших дозах способно вызвать полиорганную недостаточность и стойкую артериальную гипотонию. Подавление активности любого из них устраняет это взаимодействие и заметно улучшает состояние больного. ИЛ-1 сильнее активирует Т- и В-лимфоциты при 39*С, чем при 37*С. ИЛ-1 и ФНО альфа вызывают снижение без жировой массы тела и потерю аппетита, приводящие к кахексии при длительной лихорадке . Эти цитокины попадают в кровоток лишь на короткое время, но его оказывается достаточно, чтобы запустить продукцию ИЛ-6 . ИЛ-6 постоянно присутствует в крови, поэтому его концентрация в большей степени соответствует выраженности лихорадки и других проявлений инфекции. Тем не менее, ИЛ-6 в отличие от ИЛ-1 и ФНО альфа не считают летальным цитокином. Список использованной литературы
Аутокринно-паракринная регуляция иммунного ответа Фактор некроза опухолей
Интерлейкин 18 (интерферон-гамма-индуцирующий фактор) Интерлейкин 33 |