Элементарная структурная функциональная генетическая единица живых систем
 Скачать 104.9 Kb. Скачать 104.9 Kb.
|

|
1) Клетка – элементарная структурная функциональная генетическая единица живых систем. Роберт Гук в 1665 году структурировал первый микроскоп, он увидел пустые ячейки – целлюла (клетка), отсюда пошла цитология. Он вел термин клетки. Антонио Левенгук усовершенствовал микроскоп, он рассматривал лужу и увидел инфузорию-туфельку, открыл простейших. 1831 год – Роберт Броун открыл ядро. 1838-1839 гг. – Шванн и Шлейден сформулировали клеточную теорию. Основные положения клеточной теории Т. Шлейдена и М. Шванна (1838): 1. Все животные и растения состоят из клеток. 2. Растут и развиваются растения и животные путём возникновения новых клеток. 3. Клетка является самой маленькой единицей живого, а целый организм — это совокупность клеток. Положения современной клеточной теории: 1. Клетка – элементарная функциональная генетическая единица всех живых организмов. 2. Клетки всех одноклеточных и многоклеточных организмов сходны по строению, химическому составу, важнейшим проявлениям процессов жизнедеятельности. 3. Каждая новая клетка образуется в результате деления исходной материнской клетки. 4. Клетки специализируются и объединяются по выполняемым функциям и строению ткани. 5. Клетка является открытой системой, через которую происходят и преобразуются потоки информации, энергии и вещества. Клеточная теория позволила понять как зарождается, развивается и функционирует живой организм, а в медицине – понимания процессов жизнедеятельности и развития болезней на клеточном уровне – что открыло немыслимые ранее новые возможности диагностики, лечение заболеваний. 2) Прокариоты – это доядерные организмы, к которым относятся бактерии и синезеленые водоросли. 1. Малые размеры клеток 2. Нуклеоид – аналог ядра. Замкнутая кольцевая ДНК. 3. Отсутствуют мембранные органеллы. 4. Их функцию выполняют впячивание цитоплазматической мембраны – мезосомы. 5. Нет клеточного центра. 6. Клеткам прокариот не свойствен мейоз и митоз. Деление клеток осуществляется путем амитоза (деление интерфазного ядра путем перетяжки, без веретена деления). Одномембранные органоиды: ЭПС, Аппарат Гольджи, Лизосомы, Вакуоль Двумембранные: Ядро, Митохондрии, Пластиды, Хлоропласты. Немембранные: Микротрубочки, Клеточный центр, Реснички, Жгутики, Рибосомы. Эукариоты – ядерные организмы, к которым относятся растения, животные, грибы. Типичная эукариотическая клетка состоит из цитоплазмы, ядра, цитоплазматической мембраны. Цитоплазматический матрикс, или гиалоплазма вместе с органоидами клетки образуют цитоплазму. Матрикс обладает вязкостью, коллоидными свойствами, сократимостью, внутреннему движению. Состоит из белков, аминокислот, липидов. Цитоплазма внутренняя среда клетки, содержащая органоиды и включения. Хромопласты содержат пигменты. Лейкопласты – бесцветные пластиды, запасающая функция. Органоиды – это постоянные дифференцированные участки цитоплазмы, имеющие определённую функцию и строение. 1. Органоиды общего назначения: ЭПС, аппарат Гольджи, митохондрии, лизосомы, пластиды, клеточный центр. 2. Органоиды специального назначения: Миофибриллы (сокращение мышечных волокон), нейрофибриллы (основа нервной клетки), микроворсинки, реснички, жгутики, вакуоли у простейших. Гипотеза симбиоза и инвагинационная гипотеза. Согласно гипотезе симбиоза, первичная клетка крупной прокариотической бактерии, вступив в симбиоз с клетками сине – зелёных водорослей и приобрела пластиды. Симбиоз прокариотических бактерий с гетеротрофными прокариотическими клетками привёл их к преобразованию в митохондрии. Симбиоз со спирохетоподобными бактериями мог привести к возникновению жгутиков. По инвагинационной гипотезе, эукариотическая клетка происходит из одной аэробной прокариотической клетки (а не из многих, как в первой гипотезе). В этой предковой клетке появились впячивания клеточной мембраны, в результате продолжающейся инвагинации образовывались тельца (первичные органоиды). Отличия растительной клетки от животной: Животные клетки круглые, растительные прямоугольные, у животных клетках нет хлоропластов, есть центриоли, у растительных клетках есть центральная вакуоль. 3) а)б)в) – в альбоме г) Хромосома – это структура клеточного ядра, которая содержит гены и несет определенную наследственную информацию о признаках и свойствах организма. Состоит из первичной перетяжки – центромеры, плечей(q-длинное, p-короткое), вторичной перетяжки(отделяет от основной части), спутника, теломер (концевые участки хромосом, препятствуют слипанию и обеспечивает прикрепление к оболочке ядра). По форме различают равноплечие (метацентрические), неравноплечие (субметацентрические), палочковидные (акроцентрические). В состав хроматина входит ядерные белки (гистоновые и негистоновые) и ДНК. В начале деления хромосомы удвоены и состоят из двух дочерних клеток – хроматид. Кариотип – набор хромосом. Генотип – совокупность генов организма. Фенотип – совокупность признаков организма. Уровни упаковка хромосом: 1. Нуклеосомная нить – нить ДНК длиной 146-200 пар нуклеотидов, обернутый вокруг 4 видов гистоновых белков: H2A, H2B, H3, H4. Белки образуют белковые поры, состоящие из 8 молекул гистона (по 2 молекулы каждая). ДНК между 2 порами называется линкерная (связующая) ДНК, к ней присоединяется H1. 2. Хроматиновая фибрилла – это нуклеосомная нить, упаковывается с помощью гистона Н1 и Н2, диаметр 20-30 нм, длина 1-2 нм. 3. Интерфазная хромонема – дальнейшая упаковка хроматиновых фибрилл, белки сближаются с отдалёнными участками ДНК с образованием петли. 4. Метафазная хромонема – в метафазе каждая хромосома состоит из 2 хроматид, объединенных центромерой, к которой во время митоза прикрепляются веретена деления. В метафазе лучше всего изучать хромосомы. В анафазе хроматиды отделяются друг от друга, из них образуются дочерние хромосомы. 4) Промежуток времени от появления клетки в результате деления до ее разделения или гибели называется жизненным циклом клетки. У эукариотических клеток жизненный цикл делится на две основные стадии: интерфазу и митоз. Интерфаза — это промежуток времени в жизненном цикле, в который клетка не делится и нормально функционирует. Интерфаза делится на три периода: G1-, S- и G2-периоды. G1-период (пресинтетический, постмитотический) — это период роста и развития клетки, в который происходит активный синтез РНК, белков и других веществ, необходимых для полного жизнеобеспечения вновь образовавшейся клетки. К концу этого периода клетка может начать готовиться к удвоению ДНК. В S-периоде (синтетическом) происходит репликация ДНК. Единственным участком хромосомы, который не подвергается репликации, является центромера, поэтому образовавшиеся молекулы ДНК не расходятся полностью, а остаются скрепленными в ней, и в начале деления хромосома имеет X-образный вид. Генетическая формула клетки после удвоения ДНК — 2n4c. Также в S-периоде происходит удвоение центриолей клеточного центра. G2-период (постсинтетический, премитотический) характеризуется интенсивным синтезом РНК, белков и АТФ, необходимых для процесса деления клетки, а также разделением центриолей, митохондрий и пластид. До конца интерфазы хроматин и ядрышко остаются хорошо различимыми, целостность ядерной оболочки не нарушается, а органоиды не изменяются. Продолжительность интерфазы в клетках растений и животных составляет в среднем 10– 20 часов, тогда как митоз занимает около 1–2 часов. Генетически запрограммированная смерть клетки называется апоптозом. 5) Митоз — способ непрямого деления соматических клеток. Митоз делится на четыре основные фазы: профазу, метафазу, анафазу и телофазу. Профаза — наиболее длительная стадия митоза, в процессе которой происходит конденсация хроматина, в результате чего становятся видны X-образные хромосомы, состоящие из двух хроматид (дочерних хромосом). При этом исчезает ядрышко, центриоли расходятся к полюсам клетки, и начинает формироваться ахроматиновое веретено (веретено деления) из микротрубочек. В конце профазы ядерная оболочка распадается на отдельные пузырьки. 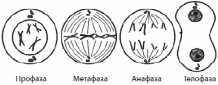 В метафазе хромосомы выстраиваются по экватору клетки своими центромерами, к которым прикрепляются микротрубочки полностью сформированного веретена деления. На этой стадии деления хромосомы наиболее уплотнены и имеют характерную форму, что позволяет изучить кариотип. В анафазе происходит быстрая репликация ДНК в центромерах, вследствие которой хромосомы расщепляются и хроматиды расходятся к полюсам клетки, растягиваемые микротрубочками. Распределение хроматид должно быть абсолютно равным, поскольку именно этот процесс обеспечивает поддержание постоянства числа хромосом в клетках организма. На стадии телофазы дочерние хромосомы собираются на полюсах, деспирализуются, вокруг них из пузырьков формируются ядерные оболочки, а во вновь образовавшихся ядрах возникают ядрышки. После деления ядра происходит деление цитоплазмы — цитокинез, в ходе которого и происходит более или менее равномерное распределение всех органоидов материнской клетки. Таким образом, в результате митоза из одной материнской клетки образуется две дочерних, каждая из которых является генетической копией материнской (2n2c). Амитозом называют прямое деление эукариотических клеток, при котором не происходит образования генетически равноценных клеток. Биологическое значение митоза – сохранение постоянства числа хромосом. 6. Размножение – способность живых организмов воспроизводить себе подобных. Существуют 2 способа размножения: бесполое и половое. Митоз обеспечивает бесполое, мейоз половое. Бесполое (шизогония(деление ядра - простейшие), почкование(грибы), вегетативное размножение(клубни картофеля), спорообразование(мох, папоротники). 7. Мейоз — это способ непрямого деления первичных половых клеток (2n2с), в результате которого образуются гаплоидные клетки (1n1с), чаще всего половые. В отличие от митоза, мейоз состоит из двух последовательных делений клетки, каждому из которых предшествует интерфаза. Первое деление мейоза (мейоз I) называется редукционным, так как при этом количество хромосом уменьшается вдвое, а второе деление (мейоз II) — эквационным, так как в его процессе количество хромосом сохраняется. ИнтерфазаI протекает подобно интерфазе митоза. Мейоз I делится на четыре фазы: профазу I, метафазу I, анафазу I и телофазу I. В профазе I происходят два важнейших процесса — конъюгация и кроссинговер. Конъюгация — это процесс слияния гомологичных (парных) хромосом по всей длине. Образовавшиеся в процессе конъюгации пары хромосом сохраняются до конца метафазы I. Кроссинговер — взаимный обмен гомологичными участками гомологичных хромосом. В результате кроссинговера хромосомы, полученные организмом от обоих родителей, приобретают новые комбинации генов, что обусловливает появление генетически разнообразного потомства. В конце профазы I, исчезает ядрышко, центриоли расходятся к полюсам клетки, а ядерная оболочка распадается. В метафазе I пары хромосом выстраиваются по экватору клетки, к их центромерам прикрепляются микротрубочки веретена деления. В анафазе I к полюсам расходятся целые гомологичные хромосомы, состоящие из двух хроматид. В телофазе I вокруг скоплений хромосом у полюсов клетки образуются ядерные оболочки, формируются ядрышки. Цитокинез I обеспечивает разделение цитоплазм дочерних клеток. Образовавшиеся в результате мейоза I дочерние клетки (1n2c). 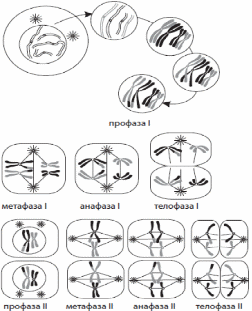 Сравнительная характеристика митоза и мейоза
Интерфаза II очень короткая, так как в ней не происходит удвоения ДНК, то есть отсутствует S-период. Мейоз II также делится на четыре фазы: профазу II, метафазу II, анафазу II и телофазу II. В профазе II протекают те же процессы, что и в профазе I, за исключением конъюгации и кроссинговера. В метафазе II хромосомы располагаются вдоль экватора клетки. В анафазе II хромосомы расщепляются в центромерах и к полюсам растягиваются уже хроматиды. В телофазе II вокруг скоплений дочерних хромосом формируются ядерные оболочки и ядрышки. После цитокинеза II генетическая формула всех четырех дочерних клеток — 1n1c, однако все они имеют различный набор генов, что является результатом кроссинговера и случайного сочетания хромосом материнского и отцовского организмов в дочерних клетках. Биологическое значение мейоза – сохранение кариотипа человека. Гаметогенез (от греч. гамете — жена, гаметес — муж и генезис — происхождение, возникновение) — это процесс образования зрелых половых клеток, яйцеклеток и сперматозоидов. Стадия созревание гаметогенеза находится в мейозе. Партеногенез – половое развитие без оплодотворения. Онтогенез – индивидуальное развитие организма. 8. Нуклеи́новые кислоты являются макромолекулами, это полимеры, состоящие из мономеров — нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание. ДНК – высокомолекулярное полимерное соединение, мономерами которого являются нуклеоитиды.Состоит из дезоксирибозы – пятиуглеродный сахар,, одноиз 4азотистых оснований (аденин, тимин, гуанин, цитозин), осток фосфорной кислоты). Соединяются нуклеотиды в полинуклеиновую цепь с помошью фосфодиэфирной связи. Сборка осуществляется ферментами полимеразой. Каждой последующий нуклеотид присоединяется к гидроксильной группе, стоящей в положении атома углерода 3 штрих. Начало цепи несёт фосфатную группу в положении 5 штрих, поэтому в нуклеотической цепи можно выделить 3 штрих и 5 штрих. Структурная организация молекулы ДНК: 1. Первичная структура – одинарная полинуклеотидная цепь. 2. Вторичная структура – 2 комплиментраные друг другу и антипараллельные полинуклеотидные цепи, соединённые водородными связями. 3. Третичная структура – трёхмерная спираль. Предложена в 1953 году Джеймсом Уотсоном и Френдсисом Криком. Диаметр спирали – 2 нм, длина ветка – 3,4 нм. Большинство молекул находятся в правозакрученной форме. Репликация – самоудвоение. В процессе репликации каждая из 2 цепей материнской молекулы ДНК служит матрицей(нуклеотидной последовательности) для дочерней, такой способ удвоения называется полуконсервативной. Материнская молекула Днк была раскручена и выкинута. С помощью фермента геликазы двойная спираль ДНК в отдельных зонах (репликонах) расплетается. Области расхождения полинуклеосомных цепей в зонах реплликаций называются репликационнымивилками. В каждой такой области с помощью фермента ДНК-полимеразы синтезируются 2 новые дочерние молекулы. В каждой репликоне ДНК-полимераза может двигаться вдоль материнской нити только в одном направлении от 5 штрих к 3 штрих концу, поэтому процесс репликации при антипараллельном соединении 2 цепей ДНК будет протекать по-разному. На одной из матриц (3 штрих к 5 штрих) сборка новой цепи идёт непрерывно от 5 штрих к 3 штрих концу. Вторая цепь ДНК осуществляет синтез короткими фрагментами также в направлении от 5 штрих к 3 штрих концу, и эти фрагменты называются фрагменты Оказаки. Фрагменты Оказаки вшиваются в единую нить ферменты лигазой из 2 синтезируемых дочерних цепей, одна строится непрерывно и её синтез идёт быстрее и называется лидирующей. 2 цепь называется запаздывающей (отстающей). Синтез медленнее. Ферменты репликации молекулы ДНК: Геликаза – делит двуцепочную мол. ДНК на 2 одноцепочные. SSB-белок – связываетет одноцепочными нитями ДНК, предотвращает комплиментарное спаривание. Топоизомераза – изменяет степень сверхспиральности, возникает при раскрученности 2 цепей ДНК. ДНК-полимераза – синтез новой цепи ДНК по принципу комплиментарности. Праймаза – образует РНК- праймеров, который синтезирует ДНК. ДНК-лигаза – образует фосфодиэфирную связь между 2 полинуклеотидами. 9) РНК – биополимер, мономеры которых является нуклеотиды. Сотоит из углевода-рибозы, азотистого основания (урацил вместо тимина). Виды РНК.Рибосомальная РНК (р-РНК) в соединении с белком входит в состав рибосом. Информационная РНК (и-РНК) синтезируется на полуцепи ДНК, несет информацию о синтезе определенного белка. Транспортная РНК (т-РНК), имеет форму трилистника (рис. 7), имеет короткую нуклеотидную последовательность. В ней выделяют четыре главные части: акцепторный стебель - одноцепочный участок, который заканчивается последовательностью нуклеотидов ЦЦА со свободной ОН группой. К этому концу присоединяется транспортируемая аминокислота, боковые петли, средняя петля антикодоновая – состоит из 5-ти нуклеотидов и содержит в центре антикодон. Антикодон - это три нуклеотида, который шифруют аминокислоту, транспортируемую к месту синтеза белка. 10) Белки - высокомолекулярные полимерные соединения мономерами, которых являются аминокислоты. Структуры белка: первичная (линейная структура), вторичная (спиральная), третичная (глобулиновая), четвертичная (результат слияния несколько глобул). Белки выполняют следующие функции: *ферментативную; * структурную; * рецепторную; * транспортную; * защитную; * двигательную; * регуляторную; * энергетическую. Строение: имеет радикал NH2 – аминогруппу, COOH – карбоксильную группу. 11) При реакциях матричного синтеза образуются полимеры, строение которых полностью определяется строением матрицы. В основе реакций матричного синтеза лежит комплементарное взаимодействие между нуклеотидами. Транскрипция (от лат. транскрипцио — переписывание) — это биосинтез молекул иРНК на матрице ДНК. Транскрипция подразделяется на три основных стадии – инициацию (начало синтеза мРНК), элонгацию (удлинение полинуклеотидной цепи) и терминацию (окончание процесса). Трансляция (от лат. трансляцио — передача) — это биосинтез полипептидной цепи на матрице иРНК, при котором происходит перевод генетической информации в последовательность аминокислот полипептидной цепи: Фаза инициации заключается в объединении двух субъединиц рибосом на определенном участке мРНК и присоединении к ней первой аминоацил тРНК. Этим задается рамка считывания информации. Фаза элонгации– тРНК с аминокислотой подходит к аминоацильному центру рибосомы, если антикодон т-РНК комплементарен кодону и РНК, то происходит временное присоединение тРНК саминокислотой к кодону иРНК. Фаза терминации.Связана с вхождением одного из трех известных стоп кадонов м РНК (УАА, УАГ, УГА) в А-участок рибосомы. Перенос генетической информации осуществляется по схеме ДНК – РНК – белок – признак. Это сложившееся представление о переносе генетической информации принято называть центральной догмой молекулярной биологии. 12) Ген – единица наследственности и изменчиваости. Это участок мол. ДНК, в котором закодирована информация о синтезе определённого полипептида. Этапы становления: 1. В 1865 году Мендель утверждал о существовании половых клетках дискретных единиц наследственных факторов, которые в 1909 году Иогансен назвал генами. 2. Томас Морган и его школа в 1908-1916 годах доказали локализацию генов в хромосомах. 3. Сложное строение гена. Серебровский и Дубинин докали, что ген делим, и внутри гена есть нуклеотиды, способные к рекомбинации (кроссинговеру) и мутации. С. Бензер: единица рекомбинации – рекон, единица мутации – мутон, цистро – соответствует фрагменту молекулы ДНК. Основные положения теории гена: 1. Ген занимает определенный участок (локус) хромосомы. 2. Ген (цистрон) - часть молекулы ДНК, имеющей определенную последовательность нуклеотидов, представляет собой функциональную единицу наследственной информации. Число нуклеотидов, входящих в состав различных генов, неодинаково. 3. Внутри гена могут происходить рекомбинации (к ней способны частицы цистрона - реконы и мутации - мутоны). 4. Существуют структурные и функциональные гены. Структурные гены кодируют синтез белков, функциональные контролируют и направляют деятельность структурных генов и в синтезе белка непосредственно не участвуют. К функциональным генам относятся: А) Ген-промотор + ген-инициатор – они указывают место начала транскрипции. Б) Ген-оператор – включает или выключает транскрипцию со структурных генов. В) Ген-терминатор – контролирует окончание транскрипции и иРНК отделяется от РНК-полимеразы. Г) Ген-регулятор - отвечает за синтез особых регуляторных белков. 5. Молекулы ДНК, входящие в состав гена, способны к репарации (восстановлению), поэтому не всякие повреждения гена ведут к мутациям. 6. Генотип (состоящий из отдельных генов) функционирует как единое целое. На функцию генов оказывают влияние факторы как внутренней, так и внешней среды. 13) Свойства гена: 1. Ген дискретен – развитие отдельного признака контролируется определённым геном. 2. Ген стабилен – как дискретная единица наследственной информации отличается постоянством и передается в поколенияхв неизменённом виде. 3. Ген лабилен – под действием факторов среды способен изменятся и мутировать. 4. Ген специфичен – каждый ген обеспечиает развитие определённого признака. 5. Ген способен к множественному плейотропномудействию – детерминирует синтез нескольких полипептидных цепей. Свойства генетического кода: 1. Триплетность – одна аминокислота соответсвует 3 нуклеотида. 2. Вырожденность – количество возможных триплетов 64 (61 кодирующие и 3 консенс кадона в ДНК АТТ, АЦТ, АТЦ, в иРНК УАА, УГА, УАГ). 3. Специфичность – каждый триплет кодирует 1 аминокислоту. 4. Неперекрываемость – 1 нуклеотид входит в состав только 1 триплета. 5. Универсальность – у всех живых организмов одинаковые триплеты кодируют одинаковую аминокислоту. 14) В альбоме. 15) Экспрессиягенов — это процесс, в ходе которого наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции. Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. У прокариот и эукариот гены представляют собой последовательности нуклеотидов ДНК. На матрице ДНК происходит транскрипция — синтез комплементарной РНК. Далее на матрице мРНК происходит трансляция — синтезируются белки. На стадии транскрипции принимают участие негенетические факторы. К ним относятся в-ва небелковой породы или эффекторы. Жакоб и Моно создали оперонную модель регуляции транскрипции. 16) При взаимодействии с белками-регуляторами они приобретают способность соединяться с геном-оператором, и если при этом транскрипция запускается, то эффекторы играют роль индукторов. Индуктор способен блокировать белок-репрессор, который не может соединяться с геном-оператором и запускается транскрипция со структурных генов. Если эффекторы препятсвуют транскрипции, их называют корепрессоры, т.к. они переводят белок-репрессор в активное состояние, он блокирует оператор и транскрипция не идёт. Триполимеразы в транскрипции: 1. РНК-полимераза1 – нах-ся в ядрышке, отв. за транскрипцию генов рРНК. 2. РНК-полимераза2 – нах-ся в ядерном соке, отв. за синтез проиРНК. 3. РНК-полимераза3 – находятся в ядерном соке, осущ-ет синтез рРНК и тРНК. Процессинг – созревание мРНК. Сплайсинг – сшивание кодирующих участков экзонов. Интроны – обеспечивают струк. организацию хроматина, запасы информаиии для изменчивости, защитная роль и действие мутагенов. Оперон – это блок тесносвязанных структурных и функциональных генов: гена-оператора, промотора, терминатора. Экзоны – информативные участки гена. Терминатор – прекращение цепи РНК. 17) Промотор (Р) – это небольшой участок ДНК перед оператором. Он служит местом связывания РНК-полимеразы. Место связывания репрессора (О) и участок Р слегка перекрываются, так что, когда репрессор находится на ДНК, РНК-полимераза не может связаться с промотором и транскрипция не идет. Энхансер – усиливают транскрипцию. 18) Особенностью регуляции транскрипции у эукариот является подчиненность этих процессов регулирующим влияниям со стороны гормонов организма. Последние часто играют роль индукторов транскрипции. Примером участия гормонов в регуляции активности определенных генов может служить влияние тестостерона на развитие тканей организма при наличии специфического белка-рецептора. Ведущая роль в компактизации ДНК принадлежит гистонам, поэтому они, несомненно, участвуют и в процессах регуляции генной активности. Условием для осуществления транскрипции у эукариот является предварительная декомпактизация хроматина на соответствующем участке, где временно утрачивается связь с H1-гистонами и несколько ослабляется связь с нуклеосомными гистонами. Возникшая в клетке потребность в каком-либо белке не может быть быстро удовлетворена путем включения транскрипции соответствующего гена. Синтезированный транскрипт должен подвергнуться процессингу, затем зрелая мРНК должна выйти из ядра в цитоплазму и, образуя комплекс с рибосомами, осуществить трансляцию информации, синтезировав пептид, который, лишь пройдя посттрансляционное изменение, формирует активный белок, необходимый клетке. В том случае, когда клетке нужно прекратить синтез какого-то продукта, после выключения транскрипции соответствующего гена вцитоплазму некоторое время будут продолжать поступать созревающие молекулы мРНК, осуществляющие там синтез пептидных цепей, пока они не деградируют под действием ферментов. Таким образом, для эффективной регуляции экспрессии генов у эукариот должны существовать механизмы, работающие не только на стадии транскрипции, но и на других этапах этого процесса. Стадии посттрансляционных изменений. Многие белки синтезируются в неактивном виде (предшественники) и после схождения с рибосом подвергаются постсинтетическим структурным модификациям. | ||||||||||||||||||||||||||||||||||||||||