В.М.Жданов. Эволюция вирусов. 1990. В.М.Жданов. Эволюция вирусов. Эволюция вирусов
 Скачать 1.63 Mb. Скачать 1.63 Mb.
|

|
ГЛАВА 2. ОСНОВНЫЕ НАПРАВЛЕНИЯ ЭВОЛЮЦИИ БИОСФЕРЫ Цель этой главы — служебная: мы постараемся напомнить читателю основные факты и заключения, относящиеся к возникновению и эволюции органической жизни на Земле, остановившись, в частности, на эволюции групп животных и рас тений, которые стали хозяевами изученных вирусов. В соответствии с современными представлениями возраст Земли определяется примерно в 4,7 (4,66) млрд лет, из которых около 1 млрд лет занимал догеологический, а 3,50— 3,57 млрд лет — геологический период. В свою очередь этот период делят на докембрий (3 млрд лет) и фенарозой (0,57— 0,585 млрд лет). Дальнейшее подразделение геологической истории Земли приведено в табл. 1. Палеоген и неоген объединяют в третичный период и в этом случае антропогену соответствует четвертичный период. Но палеоген подразделяют на эпохи (палеоцен, эоцен и олигоцен), неоген подразделяют на миоцен и плиоцен, в этом случае антропогеиу соответствует плейстоцен (1—2 млн лет). Если считать временем возникновения Земли 4—5 млрд лет назад, то в начальном периоде, длившемся 0,5 млрд лет, поверхность Земли напоминала лунный рельеф. Затем Земля перешла в первично-океаническую стадию и образовалась первичная атмосфера. Тогда же, около 4 млрд лет назад, появилась кора континентального типа. К середине и особенно к концу архейской эры (3—2,5 млрд лет назад) протоконтиненты объединились в крупные плиты, а в начале мезозоя (1,4 млрд лет назад) образовался единый материк Пангея, состоявший из северной (Лавразия) и южной (Гондвана) частей. Таблица 1. Геологическая хронология Земли
Затем, однако, произошли распад Гондваны и Лавразии (0,3 млрд лет назад) и формирование ныне существующих материков и океанов. Процесс этот был длительным, и до эоценового периода Австралия и Антарктида представляли ещё единый материк. С конца эоцена (50 млн лет назад) конфигурация континентов и океанов приобретает современный вид В истории Земли было несколько интенсивных периодов горообразования, отразившихся на эволюции биосферы. Первое великое горообразование произошло между археем и протерозоем, второе — на стыке протерозоя и кембрия. Оба они сопровождались значительным уничтожением ископаемых. В последующем были менее интенсивные горообразования которые не сопровождались большим уничтожением ископаемых: герцинское — в конце перми и начале триаса, альпийское — в конце мезозоя, в неогене — в Северной Америке. Первичные океаны составляли не более 1/10 объема нынешних, остальная их масса образовалась позже за счет дегазации внутренних частей Земли. Объем их значительно увели чился в докембрии и достиг современных размеров в кембрии Соответственно менялись рельеф и климат Земли. Архей характеризуется значительной вулканической дея тельностью, эрозией на больших площадях и слабым процессом образования осадков. На грани архея и протерозоя про изошел великий период горообразования. В это время очертания материков мало напоминали нынешние. В западном по лушарии Канада и Гренландия образовали большой материк; Бразилия и Патагония были отдельными островами. Африка была связана с Аравией и Индией; большая часть Европы была покрыта морем. Восточная Европа образовывала материк Русской платформы, отделенный от островов Азии Ураль ским проливом и морем Тетис. В протерозое, особенно в его конце, отмечаются выраженная вулканическая деятельность, многократные оледенения, а также интенсивный процесс образования осадочных пород. В кембрии, после интенсивного горообразования, наряду с высокими горами на материках преобладал низменный рельеф, а климат был умеренным. Произошло перераспределение воды и суши. Канадско-Гренландский материк сначала превращается в архипелаг, а затем глубоко погружается. В южном полушарии Южная Америка существует как единый материк, включая Мексику, Флориду и Карибскую сушу. Африка соединена с Индией, Тибетом и Сунгаро-Гобией, образуя единый материк. В Европе остается большой островной массив Финно-Сарматии. Северо-восток Азии и северо-запад Америки образуют материк Берингию. Сибирское море меняет свои очертания, выходя в Атлантику и Пацифику, а в конце кембрия превращаясь в Центрально-Азиатский пролив. В ордовике происходило значительное погружение суши и климат стал теплым даже в Антарктике. В силуре в связи с поднятием суши образовались обширные внутриконтинентальные моря, а климат стал более засушливым. В конце силура начинается каледонский горообразовательный процесс, приведший к возникновению ныне существующих Скандинавских гор, гор Казахстана, Саяно-Байкальских гор, гор Шотландии. В девоне продолжается поднятие суши, внутриконтинентальные моря обмелевают и уменьшаются, климат становится аридным, а затем наступает оледенение. Появляются пустыни и полупустыни. В карбоне снова преобладают низменные материки, климат влажный и теплый, образуются обширные болота. В конце карбона вновь происходят поднятие суши и иссушение. В перми в связи с горообразованием материки приподняты, усиливается аридность, а в южном полушарии происходит оледенение. В триасе продолжается развитие аридного климата, многие моря становятся мелководными, расширяются пустынные ландшафты. Эти же процессы продолжаются в юре, однако площадь тепловодных морей становится больше. В меловом периоде образуются многие горные системы в Европе, Азии и обеих Америках (Альпы, Гималаи, Скалистые горы, Анды), Климат становится более континентальным и прохладным. Сокращается площадь морей. В течение всего мезозоя образуются обильные отложения осадочных горных пород на всех континентах. В кайнозое климат теплеет и остается таким в течение всего палеогена. В неогене холодает, а в течение плейстоцена были 4 периода оледенения, сменявшихся периодами потепления. Последний ледниковый период окончился 25000 лет назад. Образовались, а затем исчезли сухопутные мосты между Европой и Англией, Азией и Северной Америкой, Индокитаем и Западным архипелагом. Материки приняли современное очертание. Таковы были геологические и климатические условия, в которых происходили образование и эволюция органического мира на Земле. Современные представления о происхождении жизни на Земле в значительной степени сложились под влиянием концепции А. И. Опарина (1957, 1966, 1968, 1980) и J. Haldane (1965). Отсылая читателя к этим работам, а также к трудам F. Crick (1968), М. Eigen (1971), М. Retten (1971) и других авторов, мы лишь напомним, что в соответствии с этими представлениями биологической эволюции предшествовала химическая эволюция, длившаяся более 1 млрд лет и приведшая к образованию первичных форм жизни на Земле. В происхождении и эволюции жизни выделяют три главные стадии: преджизнь, раннюю жизнь в условиях бескислородной атмосферы и позднюю жизнь, связанную с кислородной атмосферой. Первичная атмосфера Земли была водородной, она исчезла в догеологическом периоде. В результате вулканической деятельности и процессов дегазации недр Земли к началу архейской эры образовалась вторичная атмосфера. Эта атмосфера была преимущественно восстановительной и содержала водород, азот, метан, аммиак, окись углерода и воду, а также цианид водорода. Более современная точка зрения заключается в том, что состав первичной атмосферы был не восстановительный, а нитрильный (3,5 Gyr). Он включал СО2, Н2О, N2 и СО и меньше NH3, H2, CH4 и Н2. При отсутствии кислород но-озонового экрана, под влиянием электрических разрядов и ультрафиолетового облучения стало возможным образование более сложных соединений: органических кислот, аминокислот, пуриновых и пиримидиновых оснований, порфиринов [Calvin М., 1969]. Все эти вещества, образовавшиеся в атмосфере, растворялись в водах Мирового океана, постепенно насыщая их и образуя «первичный бульон». За миллионы лет он стал достаточно густым, чтобы дать начало образованию высокополимериых органических соединений.  Накоплению продуктов «первичного бульона» способствовали орогенетические циклы развития Земли — чередование геосинклинальных орогенных и посторогенных периодов. Вследствие колебаний уровней моря в местах подъема суши в мелководных бассейнах под действием испарения могла происходить концентрация органических веществ «первичного бульона». Все это ускоряло химические реакции и способствовало накоплению первичных форм жизни и ее предшественников (схема 1) [Darnell A., Doolittle К., 1986]. В экспериментах, моделирующих эти первоначальные условия нашей планеты, были получены не только аминокислоты и азотистые основания, но и их полимеры — полипептиды и полинуклеотиды. По данным S. Fox и К. Dose (1977), в этих условиях образуются протеиноиды (протобелки), включающие все 20 аминокислот с молекулярными массами порядка не скольких тысяч, которые могут обладать каталитическими свойствами. По сравнению с современной жизнью преджизнь обладала большим химическим разнообразием. Существенной ее основой явилась комплементарность пуриновых и пиримидиновых оснований, которые наряду с простыми аминокислотами (глицин, аланин, серин и пр.) образовались в первобытной атмосфере и океанах из паров аммиака, синильной кислоты, метана и двуокиси углерода. Образование полинуклеотидных цепей из пуриновых и пиримидиновых оснований создавало возможность их саморепликации с воспроизведением комплементарных полинуклеотидных цепей. Дальнейший прогресс молекулярного отбора происходил при помощи аутокатализа, в результате которого возникли более сложные молекулы типа полипептидов, нуклеиновых кислот, полисахаридов. Вначале эти процессы были независимыми, но на каком-то этапе химической эволюции независимые процессы синтеза линейных последовательностей полипептидов и нуклеиновых кислот стали взаимосвязанными, поскольку такая объединенная система катализа имела преимущество перед двумя независимыми системами в том, что скорость реакции повышалась, а продукты реакции воспроизводились с высокой точностью. Таким образом мог возникнуть первичный генетический код. Вероятно, первичный генетический код был двоичным, и отражением его является решающее значение двух первых оснований современного трехчленного генетического кода. Вначале число аминокислот, из которых формировались полипептидные цепи, не превышало 10. Такой примитивный генетический код сформировался более 3,5 млрд лет назад, и в это время появились протобионты — примитивные клеточные структуры с метаболизмом. Позже число аминокислот увеличивалось до 20, а генетический код стал трехбуквенным и избыточным (вырожденным). Универсальный генетический код формировался 3,5—2,5 млрд лет назад, и в это время появились палеокариоты с универсальным дизайном биохимических структур и их метаболизмом. Менее 2,5 млрд лет назад генетический код стал универсальным, появились прокариоты, а затем эукариоты. До последнего времени предполагалось, что каталитическими свойствами обладали только полипептиды, в то время как нуклеиновые кислоты являлись только хранителями генетической информации. Однако недавно было показано, что нуклеиновые кислоты, в частности РНК, сами по себе обладают каталитическими свойствами. Были обнаружены две группы митохондриальной РНК грибов и группа ядерной РНК простейших, у которых сплайсинг обеспечивается самими РНК (self-splicing). Это отражает три возможных эволюционных пути автокаталитических реакций. Сплайсинг нитронов группы 1 происходит в отсутствие белка и источников энергии, но требует присутствия гуанозина. При сплайсинге ядерной РНК , образуется не кольцо, а лариат. По-видимому, соответствующие интроны содержат каталитические центры или же эту функцию осуществляют так называемые ядерные РНК (И1 и И2). Во всяком случае эти свойства РНК по-новому освещают происхождение жизни. Для дальнейшего их перехода к первичным живым формам было необходимо появление матричного синтеза полипептидных цепей из имевшихся в «первичном бульоне» свободных аминокислот, а такой синтез стал возможным лишь с появлением транспортных РНК (тРНК). Каждый вид тРНК обладает двойной специфичностью — способностью узнавать определенный кодон в нуклеиновой кислоте и способностью связываться с определенной аминокислотой. Вместе с тем разные тРНК имеют общие свойства: все они состоят примерно из 80 оснований, имеют одинаковую З'-концевую последовательность ССА, сходную конфигурацию и другие свойства, обеспе чивающие возможность образования между аминокислотными остатками пептидных связей. Появление тРНК означало и по явление генетического кода, так как каждому кодону полинуклеотидной цепи теперь соответствовал определенный антикодон тРНК, а каждый вид последней связывался с определенной аминокислотой. На ранних стадиях химической эволюции репликация нуклеиновых кислот и синтез полипептидов происходили, вероятно, без органического катализа, хотя уже на этом этапе, по-видимому, существовали источники энергии типа АТФ. Однако появление полипептидов с ферментативными свойствами резко ускоряло матричный синтез полинуклеотидов и белков и знаменовало собой переход от химической эволюции к биологической [Fox S., Nakashjma Т., 1980]. К этому времени, по-видимому, относится появление белково-липидных мембран, отделявших первичные формы жизни от окружающей среды. Возможно, на этой стадии химической эволюции аутокаталитические процессы стали пространственно локализоваться на первичных мембранных белково-липидных структурах с участием полисахаридов и полинуклеотидов. Моделью таких структур являются коацерваты, в которых могли протекать гетерокаталитические реакции [Опарин А. И., Гладилин К. Л., 1980]. Здесь кончается химическая эволюция и начинается эволюция биологическая (рис. 1). В результате матричного воспроизведения соответствующих молекул в определенных условиях происходит следующее: конкуренция размножающихся молекул за вещество, энергию, пространство; вариабельность матричных молекул; воспроизведение в матричных копиях всех вариантов; естественный отбор, т. е. сохранение и размножение лишь наиболее приспособленных. Иными словами, уже на этом уровне начинают действовать законы наследственности, изменчивости и эволюции органического мира, что было хорошо иллюстрировано экспериментами, проведенными Ч. Дарвином в пробирке. 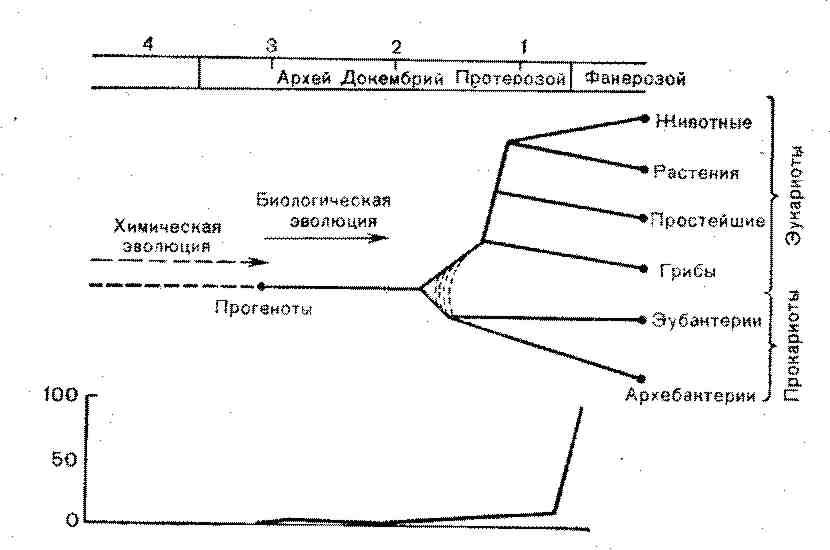 Рис. 1. Химическая и биологическая эволюция. По оси абсцисс—возраст (млрд лет); по оси ординат: вверху — эволюция органического мира, внизу—содержание кислорода в атмосфере (% к современному уровню). Для эволюции биологических макромолекул характерна самоорганизация, которая уже имеет место на уровне примитивных нуклеиновых кислот. Рассматривая вопрос, что раньше возникло, белок или нуклеиновая кислота, М. Eigen (1971) считает его некорректным, так как это все равно, что спрашивать, что возникло раньше — информация или функция. Развивая концепцию самоорганизации материи, автор выдвигает общий принцип отбора и эволюции на молекулярном уровне, основанный на критерии устойчивости стационарных состояний по нелинейной термодинамической теории. Эволюция представляется неизбежным событием, если задано присутствие определенного вещества с определенными автокаталитическими свойствами и если поддерживается такая величина порога свободной энергии, которая необходима для компенсирования стационарного производства энтропии. Эта теория дает количественную основу для постановки лабораторных экспериментов по эволюции и правила построения простых молекулярных моделей, соответствующих возможным предшественникам живых клеток. Для биологических полимеров характерна способность к самосборке. Этими свойствами, по-видимому, уже обладали предшественники биологических полимеров, образовавшихся в «первичном бульоне», в частности, протеноиды, которые легко формируют сферические (микросферы) и другие образования, характеризующиеся относительной стабильностью и определенной упорядоченностью внутренней структуры. Граничные их структуры напоминают клеточные мембраны. Вероятно, подобного рода самособранные образования и дали начало протоклетке (схема 2). В статьях А. П. Руденко (1969, 1970) и Л.Б. Меклера (1980) на пути образования протоклетки отмечены следующие процессы или стадии: возникновение упорядоченности в белках в отсутствие макромолекул и современного генетического кода; возникновение ферментов, участвующих в их синтезе; возникновение обмена веществ в отсутствие клеток, «наделенных» обменом; возникновение клеток в отсутствие других клеток, которые могли дать им начало; возникновение систем аккумулирования энергии; возникновение генетического кода. Предполагают, что первичные формы жизни содержали в качестве генетического материала РНК. ДНК появилась позже и с ее появления произошло разделение функций между ДНК и РНК в том виде, в котором они существуют в настоящее время: ДНК является хранителем генетической информации, а из двух видов РНК мРНК кодирует генетическую информацию с ДНК, а тРНК переводит генетический код с языка нуклеинового на язык белковый. Появление рибосомной РНК (рРНК) и возникновение рибосом целиком относятся к ранним этапам биологической эволюции, и все сохранившиеся до нашего времени древнейшие формы жизни обладают рибосомными системами синтеза белков. Стабильность структуры тРНК и рРНК на протяжении нескольких миллиардов лет эволюции свидетельствует об исключительно стабилизирующем характере естественного отбора в этом случае. Предполагают, что переход от протоклетки (proto-cell) к прокариотной клетке произошел в промежутке 3—4 млрд лет назад. Предшественниками ископаемых бактерий были прогеноты. Ранние прокариотические клетки существовали в восстановительной атмосфере и нынешние метаногенетические бактерии, по-видимому, являются прямыми их потомками. Первоначальные прокариоты обладали способностью к анаэробному брожению. Затем появилась примитивная форма бактериального фотосинтеза, затем более сложная форма фотосинтеза типа растений (сине-зеленых водорослей) и лишь впоследствии аэробное дыхание. Образования, проходившие долгий путь от неживой к живой материи, обозначаются как эобионты, более близкие предшественники ископаемых бактерий (более 3 млрд лет назад), как протоклетки, или прогеноты. По данным S. Fox и соавт. (1977), изучавших гомологии 18S рРНК в целях исследования филогенеза прокариотов, первоначальные прокариоты дали начало трем эволюционным линиям: уркариотам (Urkaryotes), ставшим симбионтами, потомками которых являются цитоплазматические компоненты эукариотов; архебактерии (Archaebacteria), потомками которых являются современные метанобактерии, и эубактерии (Eubacteria) —предки современных бактерий. Схема 2. ЭВОЛЮЦИЯ САМОСБОРКИ |