В.М.Жданов. Эволюция вирусов. 1990. В.М.Жданов. Эволюция вирусов. Эволюция вирусов
 Скачать 1.63 Mb. Скачать 1.63 Mb.
|

|
ГЛАВА 3. КЛАССИФИКАЦИЯ ВИРУСОВ Прежде чем излагать данные об эволюции разных групп вирусов, целесообразно сопоставить существующую классификацию с некоторыми соображениями о возможных источниках происхождения и об основных направлениях эволюции вирусов. Использовав основные критерии международной классификации вирусов, мы построили ее по принципу «от простого — к сложному» (в скобках указаны порядковые номера). A. Прионы Б. Вироиды B. РНК-содержащие вирусы 1. РНК однонитевая 1.1. Без оболочек 1.1.1. Монопартитные 1.1.1.1. Изометрические Picornaviridae, Caliciviridae, Nudaurelia B, Leviviridae, MCDV, Tymovirus, Luteovirus, Tombusvirus, Sobemovirus, Necrovirus (1—10) 1.1.1.2. Палочковидные Closterovirus, Carlavirus, Potzvirus, Potexvirus. Tebamovirus (11—15) 1.1.2. Бипартитные 1.1.2.1. Изометричные Dianthovirus, Comovirus, Nepovirus, PEMV, Nodaviridae, VTM (21) 1.2.2.2. Палочковидные Tobravirus (22) 1.1.3. Мультипартитные 1.1.3.1. Изометрические Cucumovirus, Bromovirus, Harvirus (23—25) 1.1.3.2. Палочковидные Hordeivirus (26) 1.1.3.3. Смешанные ALMV (27) 1.2. С оболочками 1.2.1. Без синтеза ДНК 1.2.1.1. Геном позитивный Togaviridae, Flaviviridae, Coronaviridae, TSWV (28—31) 1.2.1.2. Геном негативный 1.2.1.2.1. Геном непрерывный Paramyxoviridae, Rhabdoviridae (32—33) 1.2.1.2.2. Геном фрагментарный Orthomyxoviridae, Bremyaviridae, Arenaviridae (34—36) 1.2.2. Синтез ДНК Retroviridae (37) 2. РНК двунитевая 2.1. Без оболочек 2.1.1. РНК непрерывная Totiviridae (38) 2.1.2. РНК бисегментная Partitiviridae, Birnaviridae (31—40) 2.1.3. РНК трисегментная Трисегментные миковирусы (41) 2.1.4. РНК мультисегментная Reoviridae (42) 2.2. С оболочками Cystoviridae (43) Г. Плазмиды Д. ДНК-содержащие вирусы 1. ДНК однонитевая 1.1. Без оболочек 1.1.1. Изометрические Mycroviridae Parvoviridae (44—45) 1.1.2. Пулевидные Фаги микоплазм (46) 1.2. С оболочками Plasmaviridae (47) 2. ДНК двунитевая 2.1. Без оболочек 2.1.1. Монопартитные Papovaviridae, Adenoviridae, Iridoviridae, Myoviridae, Stylovirldae, Podoviridae, Tectoviridae (48—54) 1.2.1. Мультипартитные Polydnaviridae (55) 2.2. С оболочками Plasmaviridae, Hepadnauiridae, Baculoviridae, Herpesviridae (56—59) 2.3. Co сложным строением Poxviridae (60) Мы привели перечень семейств, родов и групп вирусов из последнего издания классификации и номенклатуры вирусов [Matthews R., 1982], сгруппировав их несколько по-иному (по степени возрастания сложности) и дополнив позже предложенными таксономическими группами. Нетрудно видеть, что международная классификация вирусов, будучи вполне удобной для практических целей, далеко не отражает возможных направлений эволюции вирусов и разные ее разделы построены на различных принципах. После типа РНК (однонитевая, двунитевая) следующим по значению критерием является наличие внешних оболочек, затем моно- или мультипартитность для необолочечных вирусов (все они имеют позитивный геном) и стратегия генома для оболочечных вирусов. Нам представляется важным построить классификацию по несколько иным принципам, взяв в качестве основного критерия последнее, а именно стратегию вирусного генома. Однако, прежде чем классифицировать по этому признаку, сформулированному главным образом для вирусов животных [Baltimore D., 1974], нам хотелось бы обратить внимание на круг хозяев, поражаемых разными грунтами РНК-содержащих вирусов. Среди хозяев вирусов с двунитевой РНК находятся бактерии, грибы, растения, беспозвоночные и позвоночные животные, среди хозяев вирусов с однонитевой ДНК — одна весьма специализированная группа бактериальных вирусов, а затем многочисленные вирусы растений и животных (преимущественно высших позвоночных), а также сравнительно поздно появившихся беспозвоночных (преимущественно насекомых). Особенностью РНК-содержащих вирусов, за исключением ретровирусов, является наличие РНК-зависимой РНК-полимеразы. Этот фермент отсутствует у животных, но имеется у растений. По-видимому, растения явились источником проис хождения многих РНК-содержащих вирусов, так как РНК-зависимые синтезы РНК имеют место в растительных клетках. В пользу этого предположения косвенно свидетельствует также сложное строение некоторых репликаз у вирусов растений. Так, репликаза РНК-содержащего вируса желтой мозаики турнепса состоит из двух субъединиц, из которых одна (молекулярная масса 150000) кодируется вирусным геномом, а другая (молекулярная масса 45000) —геномом клеток растения - хозяина вируса. У растений имеется РНК-зависимая РНК-полимераза, однако некоторые вирусы растений, используя этот «хозяйский» компонент, обладают собственной полимеразой как дополнительным и единственным ферментом. В случае вируса желтой мозаики турнепса, относящегося к тимовирусам, «хозяйский» компонент (молекулярная масса 45000) и вирусный компонент (молекулярная масса 115000) необходимы для функционирования довольно простого генома этого вируса. Впрочем, другими источниками происхождения РНК-содержащих вирусов могли стать и грибы (у которых, по-видимому, существуют системы РНК-зависимого синтеза РНК), а также бактерии, простейшие, вирусы которых, будучи специализированными формами, содержат в составе репликазы также вирусный и «хозяйский» компоненты. К вопросу о происхождении и дальнейшей эволюции РНК-содержащих вирусов мы еще вернемся при рассмотрении отдельных их групп, здесь же уместно обратить внимание на разные пути совершенствования их геномов. Наиболее простыми вирусами являются группы растительных вирусов с небольшим позитивным геномом, у которых отсутствуют специальные структуры на 5'- и З'-концах молекулы РНК. Обозначим их условно первой группой с подразделением на изометрические и палочковидные. 1. Простые вирусы: а) изометрические — Nudaurelia b, Maize chlorotic dwarf virus group (MCDV), Luteovirus, Dianthovirus, Leviviridae; б) палочковидные — Closterovirus, Carlavirus. К сожалению, мы мало знаем о репликации этих вирусов, и возможно, определение «простые» является ошибочным. Поэтому лучше эти группы вирусов обозначить как малоизученные. Зато мы знаем, в каких направлениях шло совершенствование их генома. Геном любого, даже самого примитивного вируса имеет не менее 3—4 генов, если, конечно, этот вирус недефектен, как дельта-вирус — «спутник» вируса гепатита В. На нем закодированы белки капсида и полимераза, которые не должны быть синтезированы в одинаковом числе копий. Поэтому должен существовать какой-то механизм регуляции этого синтеза. На примере более изученных левивирусов (фаги QP, MS2) было показано, что эта задача решается двумя путями: расположением генов на молекуле РНК и регуляцией их с помощью синтезированных вирусных белков. Именно поэтому у столь примитивных вирусов (Leviviridae) синтез их 4 белков — белка капсида, белка созревания (белок А), литического белка и полимеразы — строго регулирован, поскольку на один зрелый вирион требуется 180 копий капсидного белка, одна копия белка А, вероятно, единичные молекулы литического белка и полимеразы. Последняя, как и у тимовируса (вируса желтой мозаики турнепса), имеет субъединичное строение, одним компонентом которого является вирусный белок, а другим — «хозяйский» белок. После того как полимераза синтезирована, она блокирует работу генов, синтезирующих уникальные белки, и этим обеспечивается многократное функционирование гена капсидного белка. Возможно, по этому же пути происходит регулирование геномов рассматриваемых двух групп простейших вирусов. В дальнейшем геном РНК-содержащих вирусов совершенствовался в нескольких направлениях. 1. Формирование структур, характерных для мРНК эукариотов: кэп-структур на 5'-концах и поли (А)-последовательностей на З'-концах, которые у вирусов растений чаще заменены структурами, сходными с тРНК и даже обладающими аминоацилакцептирующей активностью,— Tytnovirus (икосаэдрический), Tobamouirus (палочкообразный), Potexvirus (нитеобразный с кэп-структурой, но без тРНК), Necrovirus (ико саэдрический квазикэпобразная структура). 2. Синтез субгеномных РНК, кодирующих группы белков с последующим нарезанием конечных продуктов: а) Tombusvirus — икосаэдрический, к ним примыкают рассмотренные выше группы Tymovirus, Potexvirus, Tobamovirus, имеющие кэп-структуры; б) Togaviridae, Flaviviridae, Coronaviridae —все они являются оболочечными вирусами с кубическим типом симметрии нуклеокапсидов. 3. Разделение генома на сегменты, размещенные в одном и том же вирионе. Tomato spotted wiltvirus (TSWV) — оболочечный вирус со спиральным типом симметрии внутреннего рибонуклеопротеида. 4. Разделение генома на сегменты и размещение их в разных частицах (мультипартитность): а) простые бипартитные вирусы — Nodeviridae, Velvet tobacco mosaic virus group (VTMV)—икосаэдрические, Ilarvirus — палочковидные; б) вирусы со структурами кэп-тРНК — Cucumovirus, Bromovirus, Hordeivirus (кэп-тРНК-структуры), Tobravtrus, Alfalfa mosaicvirus group (ALMV) (только кэп) —палочковидные. 5. Вирусы со структурой VPg-pA: а) геном считывается в виде одного полипротеида, который затем нарезается клеточными и вирусными протеазами,— Picornaviridae (икосаэдрические), Potyvirus (нитевидные); б) синтезируются субгеномные РНК—Caliciviridae, Sobemovirus (икосаэдрические); в) субгеномные РНК разобщены в разных частицах — Соmovirus, Nepovirus, Peaenation mosaic virus (PEMV) —бипартитиые икосаэдрические. 6. Вирусы с негативно-полярным геномом: а) РНК непрерывная, гены считываются отдельно — Rhabdoviridae, Paramyxoviridae; б) РНК фрагментарная — Bunyaviridae, Orthomyxoviridae. 7. Вирусы с амбисенс-РНК — Arenaviridae. 8. Вирусы с обратной транскрипцией — Retroviridae 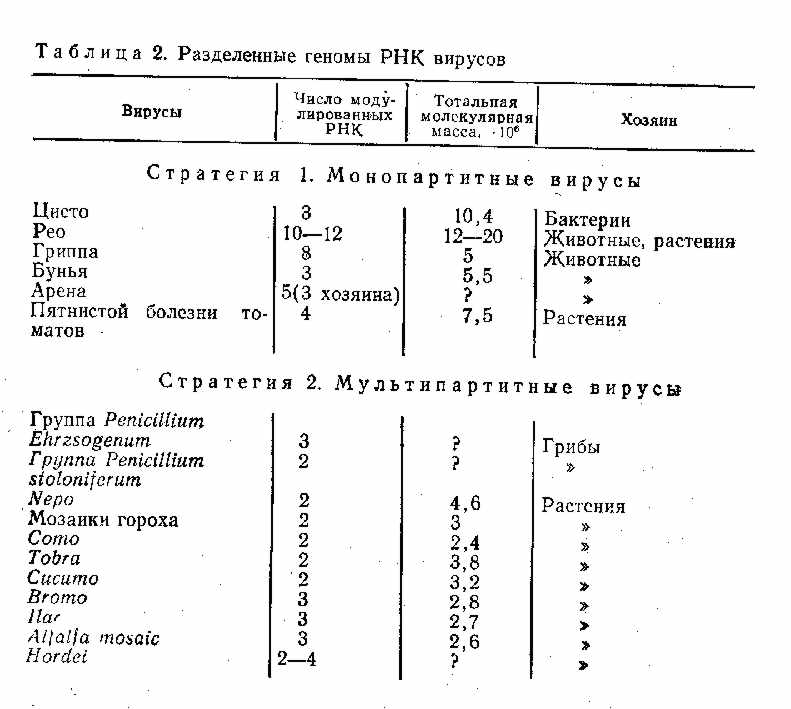 Естественно, что у некоторых вирусов различные направления эволюции «переплетаются». Таковы колицивирусы, у которых наряду со структурой VPg синтезируются субгеномные РНК, комовирусы, у которых наряду со структурой VPg имеет место бипартитность, кукумовирусы, у которых наряду с бипартитностью РНК имеются кэп-структуры и т. п. Поэтому при описании отдельных групп вирусов мы вынуждены делать отступления как от международной классификации, так и от строгого подразделения стратегий реализации генетической информации. 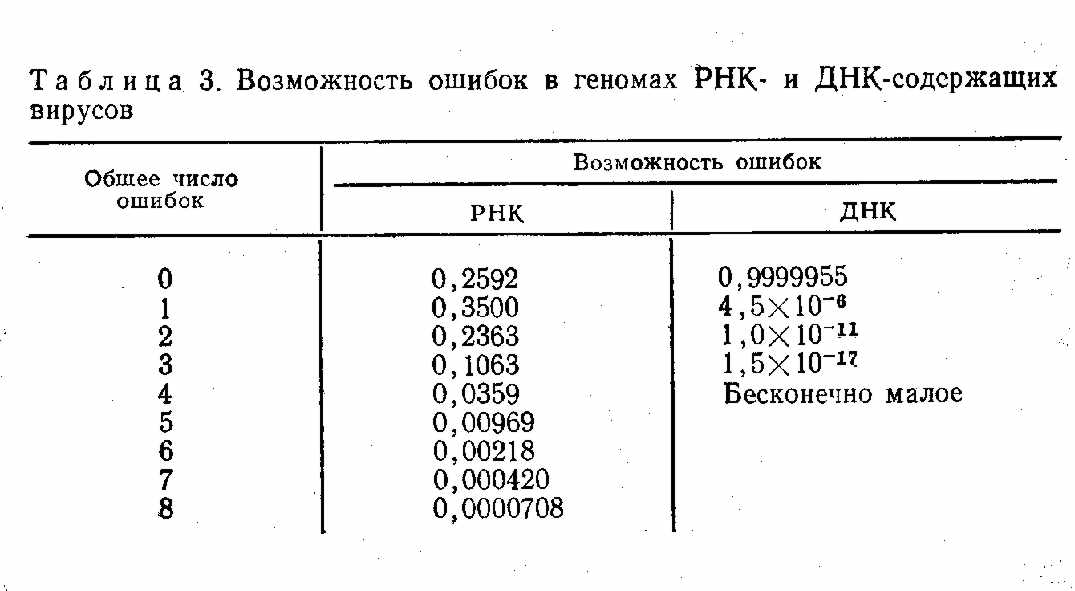 Вопрос о мультисегментности и мультипартитности вирусов явился предметом специального рассмотрения [Reanney D., 1984]. С одной стороны, мультисегментность и мультипартитность обеспечивают сбалансированный синтез отдельных белков. Однако при летальных мутациях в первом случае достаточно повредить один ген, чтобы весь нирион перестал быть инфекционным. Во втором случае летальная мутащия затрагивает только пораженный ген и не сказывается на всей популяции. С другой стороны, мультипартитные вирусы обеспечивают инфекцию лишь при достаточно массовых дозах заражающего вируса. С этими соображениями приходится считаться при оценке вероятности инфекции у животных и растений (табл. 2). Пути эволюции будут рассмотрены в разделах, посвященных разным группам вирусов. Здесь же отметим, что стратегия геномов, впервые сформулированная Reanney и Baltimore (1974), нашла дальнейшее развитие в работах В. И. Агола (1978), А. Д. Альтштейна и Н. В. Каверина (1980). А. Д. Альтштейн и Н. В. Каверин (1980) выделяют 6 стратегий реализации генетической информации. 1. (±) ДНК --> РНК --> белок (вирусы с двунитевой ДНК). 2. ( + ) ДНК --> (±) ДНК -->РНК --> белок (вирусы с однонитевой ДНК). 3. ( + ) РНК --> ДНК --> (+) РНК --> белок (ретровирусы). 4. ( + ) РНК --> (—) РНК --> (+) РНК --> белок (пикорнавирусы). 5. (—) РНК--> (+) РНК --> белок (вирусы с негативным геномом). 6. (±) РНК --> (+) РНК --> белок (реовирусы), Рассматривая происхождение вирусов, авторы считают их происходящими из первичных генетических систем репликации —-трансляции. Иными словами, вирусные генетические системы существовали с момента возникновения первой биологической генетической системы, которая по простоте и поведению соответствовала вирусным системам. Авторы считают, что появление принципиально новой вирусной системы — чрезвычайно редкое событие и большинство известных в настоящее время вирусов являются результатом эволюции генетических систем, появившихся первично или обособившихся от клеточного генома. Необходимо также отметить, что частота ошибок считывания РНК в 100000—10000000 выше частоты считывания ДНК [Reanney D., 1984] (табл. 3). Все эти данные будут учтены при рассмотрении эволюции разных групп вирусов. |