5 полный ответ. Генерация потенциала действия
 Скачать 146.38 Kb. Скачать 146.38 Kb.
|

|
Генерация потенциала действия Все клетки возбудимых тканей при действии различных раздражителей достаточной силы способны переходить в состояние возбуждения. К возбудимым относятся нервная, мышечная и железистая ткани, хотя четкой границы между возбудимыми и другими тканями провести нельзя. Возбудимость — это способность клеток к быстрому ответу на раздражение, проявляющемуся через совокупность физических, физико-химических процессов и функциональных изменений. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Опыт показывает, что возбужденный участок клетки становится электроотрицательным по отношению к невозбужденному, что указывает на перераспределение ионов в возбужденном участке. При возбуждении оно имеет временный характер, и после окончания возбуждения вновь восстанавливается исходный потенциал покоя. Потенциал действия — это сдвиг мембранного потенциала, возникающий в ткани при действии порогового и сверхпорогового раздражителя, что сопровождается перезарядкой клеточной мембраны. При действии порогового или сверхпорогового раздражителя изменяется проницаемость клеточной мембраны для ионов в различной степени. Для ионов Na она повышается в 400—500 раз, и градиент нарастает быстро, для ионов К — в 10—15 раз, и градиент развивается медленно. В результате движение ионов Na происходит внутрь клетки, ионы К двигаются из клетки, что приводит к перезарядке клеточной мембраны. Наружная поверхность мембраны несет отрицательный заряд, внутренняя — положительный.На кривой потенциала действия выделяют следующие фазы: 1. Локальный ответ (местная деполяризация), предшествующий развитию ПД. 2. Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше нуля. Поэтому мембрана приобретает противоположный заряд – внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мс. 3. Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мс. 4. Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мс. 5. Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мс. Амплитуда ПД скелетных мышц в среднем 120-130 мВ, нейронов 80-90 мВ, гладкомышечных клеток 40-50 мВ. Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые, и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных ворот зависит величина КУД (чем она выше, тем ниже КУД, и наоборот). Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ), сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию. Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой. Следовая гиперполяризация связана с повышенной после генерации ПД калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос, выносящий вошедшие в клетку во время ПД ионы натрия. Изменяя проводимость быстрых натриевых и калиевых каналов, можно влиять на генерацию ПД, а, следовательно, на возбуждение клеток. При полной блокаде натриевых каналов, например, ядом рыбы тетродонта – тетродотоксином, клетка становится невозбудимой. Это используется в клинике. Такие местные анестетики, как новокаин, дикаин, лидокаин тормозят переход натриевых каналов нервных волокон в открытое состояние. Поэтому проведение нервных импульсов по чувствительным нервам прекращается, наступает обезболивание (анестезия) органа. При блокаде калиевых каналов затрудняется выход ионов калия из цитоплазмы на наружную поверхность мембраны, т.е. восстановление МП. Поэтому удлиняется фаза реполяризации. Этот эффект блокаторов калиевых каналов также используется в клинической практике. Например, один из них хинидин, удлиняя фазу реполяризации кардиомиоцитов, урежает сердечные сокращения и нормализует сердечный ритм. Также следует отметить, что чем выше скорость распространения ПД по мембране клетки или ткани, тем выше ее проводимость. Критический уровень деполяризации Условием возникновения потенциала действия является критическая деполяризация мембраны. Если исходный потенциал мембраны перед нанесением раздражающего стимула обозначить Ео, а критическую величину мембранного потенциала EK, то указанное условие порогового раздражения можно записать так: Eo+V=Eк, где V—пороговый потенциал, т.е. величина, на которую необходимо повысить внутренний потенциал мембраны для возникновения импульса. Эта простая зависимость помогла понять причину известных со времени классических исследований Пфлюгера (1859) изменений возбудимости нервного волокна в области катода и анода постоянного тока. В области катода мембрана деполяризуется, т. е. значение Ео приближается к величине Ек, соответственно V уменьшается и, следовательно, возбудимость возрастает. В области анода, напротив, Ео уменьшается (внутренний потенциал мембраны становится более отрицательным); теперь для критической деполяризации мембраны необходимо ее потенциал сместить на большую величину V—возбудимость снижается. При длительной деполяризации мембраны развиваются процессы, повышающие критический уровень деполяризации. Такими процессами являются инактивация натриевых каналов и активация калиевых. Рост Ек при данном значении Ео ведет к увеличению порогового потенциала, т.е. снижению возбудимости. Одновременно с увеличением порога происходит снижение потенциала действия — крутизна его нарастания и амплитуда падают. Такое снижение возбудимости нервного волокна при длительной и сильной деполяризации мембраны было впервые описано Вериго (1889) и получило название катодической депрессии. Катодическая депрессия – снижение возбудимости и проводимости под катодом при длительном действии постоянного тока. Анодическая экзальтация – повышение возбудимости и проводимости под анодом при длительном действии постоянного тока. Катэлектротон – повышение возбудимости и проводимости под катодом при действии постоянного тока. Анэлектротон – снижение возбудимости и проводимости под анодом при действии постоянного тока. Порог возбудимости Порог возбудимости – это минимальная сила раздражителя, которая способна вызвать процесс возбуждения. Раздражители меньшей или большей силы называют соответственно – подпороговыми и сверхпороговыми. Порог возбуждения нерва ниже, чем порог возбуждения мышцы и особенно железы. Состояние ткани (работа, утомление, уровень метаболизма) также влияют на величину порога. Признаки возбуждения определяются формой перехода от состояния покоя к деятельности, так для нервной ткани – это генерация распространяющегося нервного импульса, синтез и разрушение медиаторов, для мышечной – сокращение, для железистой – образование и выделение секрета. Возбуждение может быть местным и распространяющимся. Законы раздражения возбудимых тканей Законы раздражения отражают определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани. Закон силы: чем больше сила раздражителя, тем больше величина ответной реакции. В соответствии с этим законом функционируют сложные структуры, например, скелетная мышца. Амплитуда ее сокращений от минимальных (пороговых) величин постепенно увеличивается с увеличением силы раздражителя до субмаксимальных и максимальных значений. Это обусловлено тем, что скелетная мышца состоит из множества мышечных волокон, имеющих различную возбудимость. Поэтому на пороговые раздражители отвечают только те мышечные волокна, которые имеют самую высокую возбудимость, амплитуд, мышечного сокращения при этом минимальна. С увеличением силы раздражителя в реакцию вовлекается все большее количество мышечных волокон, и амплитуда сокращения мышц все время увеличивается. Когда в реакцию вовлечены все мышечные волокна, составляющие данную мышцу, дальнейшее увеличение силы раздражителя не приводит к увеличению амплитуды сокращения. Закон «все или ничего»: подпороговые раздражители не вызывают ответной реакции ("ничего"), на пороговые раздражители возникает максимальная ответная реакция ("все"). По закону "все или ничего' сокращаются сердечная мышца и одиночное мышечное волокно. Закон "все или ничего" не абсолютен. Во-первых, на раздражители подпороговой силы не возникает видимой ответной реакции, но в ткани происходят изменения мембранного потенциала покоя в виде возникновения местного возбуждения (локального ответа). Во-вторых, сердечная мышца, растянутая кровью, при наполнении ею камер сердца, реагирует по закону "все или ничего", но амплитуда ее сокращений будет больше по сравнению с сокращением сердечной мышцы, не растянутой кровью. Закон силы раздражения устанавливает зависимость ответной реакции от силы раздражителя. Эта зависимость неодинакова для отдельных клеток и для целой ткани. Для одиночных клеток зависимость называется «все или ничего». Характер ответной реакции зависит от достаточной пороговой величины раздражителя. При воздействии подпороговой величиной раздражения ответной реакции возникать не будет (ничего). При достижении раздражения пороговой величины возникает ответная реакция, она будет одинакова при действии пороговой и любой сверхпороговой величины раздражителя (часть закона – все). Для совокупности клеток (для ткани) эта зависимость иная, ответная реакция ткани прямо пропорциональна до определенного предела силе наносимого раздражения. Увеличение ответной реакции связано с тем, что увеличивается количество структур, вовлекающихся в ответную реакцию. Закон раздражения - Дюбуа-Реймона (аккомодации): раздражающее действие постоянного тока зависит не только от абсолютной величины силы тока или его плотности, но и от скорости нарастания тока во времени. Закон градиента раздражения. Градиент – это крутизна нарастания раздражения. Ответная реакция ткани зависит до определенного предела от градиента раздражения. При сильном раздражителе примерно на третий раз нанесения раздражения ответная реакция возникает быстрее, так как она имеет более сильный градиент. Если постепенно увеличивать порог раздражения, то в ткани возникает явление аккомодации. Аккомодация – это приспособление ткани к медленно нарастающему по силе раздражителю. Это явление связано с быстрым развитием инактивации Na-каналов. Постепенно происходит увеличение порога раздражения, и раздражитель всегда остается подпороговым, т. е. порог раздражения увеличивается. Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает. Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого - окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще. Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка. Закон силы-времени: раздражающее действие постоянного тока зависит не только от его величины, но и от времени, в течение которого он действует. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения. Закон длительности раздражений. Ответная реакция ткани зависит от длительности раздражения, но осуществляется в определенных пределах и носит прямо пропорциональный характер. Существует зависимость между силой раздражения и временем его действия. Эта зависимость выражается в виде кривой силы и времени. Эта кривая называется кривой Гоорвега—Вейса—Лапика. Кривая показывает, что каким бы сильным ни был бы раздражитель, он должен действовать определенный период времени. Если временной отрезок маленький, то ответная реакция не возникает (как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют). Причиной такой зависимости является мембранная емкость. Очень "короткие" токи просто не успевают разрядить эту емкость до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой (минимальной силой раздражения, которая вызывает первичную ответную реакцию). Время, в течение которого действует ток, равный реобазе, называется полезным временем. В связи с тем, что определение этого времени затруднено, было введено понятие хронаксия - минимальное время, в течение которого ток, равный двум реобазам, должен действовать на ткань, чтобы вызвать ответную реакцию. Определение хронаксии - хронаксиметрия - находит применение в клинике. Электрический ток, приложенный к мышце, проходит через как мышечные, так и нервные волокна и их окончания, находящиеся в этой мышце. Так как хронаксия нервных волокон значительно меньше хронаксии мышечных волокон, то при исследовании хронаксии мышцы практически получают хронаксию нервных волокон. Если нерв поврежден или произошла гибель соответствующих мотонейронов спинного мозга (это имеет место при полиомиелите и некоторых других заболеваниях), то поисходит перерождение нервных волокон и тогда определяется хронаксия уже мышечных волокон, которая имеет большую величину, чем нервных волокон. Закон полярного действия постоянного тока: при замыкании тока возбуждение возникает под катодом, а при размыкании - под анодом. Прохождение постоянного электрического тока через нервное или мышечное волокно вызывает изменение мембранного потенциала покоя. Так, в области приложения к возбудимой ткани катода положительный потенциал на наружной стороне мембраны уменьшается, возникает деполяризация, которая быстро достигает критического уровня и вызывает возбуждение. В области же приложения анода положительный потенциал на наружной стороне мембраны возрастает, происходит гиперполяризация мембраны и возбуждение не возникает. Но при этом под анодом критический уровень деполяризации смещается к уровню потенциала покоя. Поэтому при размыкании цепи тока гиперполяризация на мембране исчезает, и потенциал покоя, возвращаясь к исходной величине, достигает смещенного критического уровня и возникает возбуждение. Закон физиологического электротона: действие постоянного тока на ткань сопровождается изменением ее возбудимости. При прохождении постоянного тока через нерв или мышцу порог раздражения под катодом и соседних с ним участках понижается вследствие деполяризации мембраны - возбудимость повышается. В области приложения анода происходит повышение порога раздражения, т. е. снижение возбудимости вследствие гиперполяризации мембраны. Эти изменения возбудимости под катодом и анодом получили название электротона (электротоническое изменение возбудимости). При дальнейшем действии постоянного тока первоначальное повышение возбудимости под катодом сменяется ее понижением, развивается так называемая католическая депрессия. Первоначальное же снижение возбудимости под анодом сменяется ее повышением анодная экзальтация. При этом в области приложения катода происходит инактивация натриевых каналов, а в области действия анода происходит снижение калиевой проницаемости и ослабление исходной инактивации натриевой проницаемости. Хронаския, лабильность Хронаксия - это наименьшее время, в течение которого электрический ток, равный удвоенной реобазе, должен действовать на ткань, чтобы вызвать возбуждение. Полезное время и хронаксия характеризуют скорость возникновения возбуждения при действии раздражителя. Для определения хронаксии применяются специальные приборы - хронаксиметры. Определение хронаксии - хронаксиметрия - получило распространение не только в эксперименте, но и в клинической практике (А. Бургиньон, Ю. М. Уфлянд и др.). В частности, путем измерения хронаксии мышцы невропатолог может установить наличие повреждения волокон двигательного нерва. Дело в том, что при приложении электрического стимула к мышце ток проходит и через находящиеся в ней нервные волокна и их окончания. Порог раздражения и хронаксия нервных волокон ниже, чем мышечных волокон. Поэтому при раздражении мышцы возбуждение прежде возникает в нервных волокнах и от них уже передается мышечным волокнам. Из этого следует, что при определении хронаксии нормальной мышцы человека фактически измеряется хронаксия иннервирующих ее нервных волокон. Если же нерв поврежден или произошла гибель иннервирующих мышцу клеток в спинном мозгу, то нервные волокна перерождаются, и тогда приложенный к мышце стимул выявляет хронаксию мышечных волокон, которая имеет большую продолжительность. Функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях. Понятие «лабильность» введено русским физиологом Н. Е. Введенским (1886), который считал мерой лабильности наибольшую частоту раздражения ткани, воспроизводимую ею без преобразования ритма. Лабильность – это свойство, отражающее функциональную подвижность возбудимых тканей. Данное понятие предложено известным Российским физиологом, учеником И.М.Сеченова, Н.Е.Введенским. По определению Н.Е.Введенского лабильность – это «большая или меньшая скорость тех элементарных реакций, которыми сопровождается физиологическая деятельность данного аппарата». Мерой лабильности является максимально возможное число элементарных циклов возбуждения, которое может воспроизвести возбудимая ткань в единицу времени в соответствии с частотой предъявленного раздражителя. Если частота раздражителя превысит меру лабильности возбудимой ткани, в последней возникнет феномен торможения. Торможение в этом случае будет выполнять охранительно-восстановительную функцию. В качестве примера такого повышения лабильности, или, как его называл А.А. Ухтомский, усвоения ритма, приведем следующий опыт Г. Мевеса на изолированном нервном волокне лягушки. Одиночное нервное волокно раздражали ритмическими стимулами частотой 460 в секунду. На каждый стимул возникал потенциал действия. Затем повышали частоту стимуляции до 740 в секунду. Вначале волокно отвечало только на каждый второй стимул, т. е. происходила трансформация ритма раздражений. Однако после нескольких секунд такого раздражения волокно начало усваивать навязанный ему ритм, и частота ответов повысилась до 740 импульсов в секунду. Усвоение ритма связано с убыстрением тех процессов обмена веществ, которые обеспечивают активное «выкачивание» из протоплазмы в наружный раствор ионов Nа, проникших через мембрану во время возбуждения. 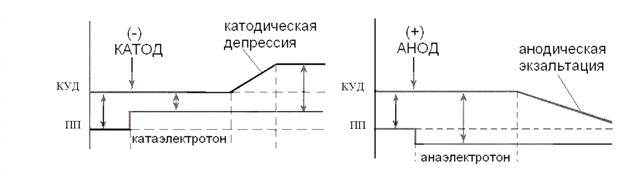 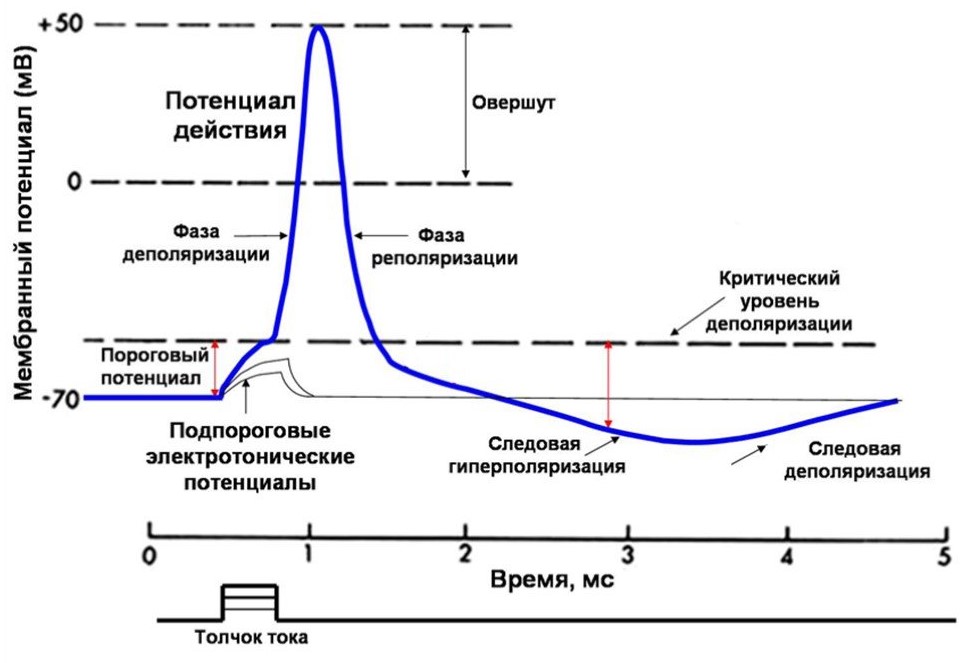 |