Министерство образования и науки Российской Федерации
Федеральное государственное бюджетное образовательное учреждение
Высшего профессионального образования
Курганский государственный университет (КГУ)
Кафедра общей и социальной психологии
Контрольная работа по дисциплине «История»
на тему:
«Homo sapiens: основные антропологические характеристики эпохи праобщины»
Выполнила студентка 1 курса
группы ПСВ-103.18
Максимова А.Н.
Проверил кандидат
исторических наук, доцент
Федоров С.Г.
2018 год
Содержание
Введение……………………………………………………………...………..3
Основная часть………………………………………………………………..4
Homo sapiens…………………………………………………………………..5
Антропогенез………...………………………………………………………..7
Антропологические особенности Homo sapiens эпохи праобщины……....9
Социологические особенности Homo sapiens эпохи праобщины.……….13
Концепции моноцентризма и полицентризма……………………………..16
Заключение…………………………………………………………………..18
Список литературы………………………………………………………….19
Введение
Специальной наукой о человеке как особом биологическом виде является антро�пология. По сравнению с прошлым столетием, когда с антропологией связывали весь комплекс научных знаний о человеке, современная антропология значительно огра�ничила свой предмет проблемами антропогенеза, расогенеза и изучением «вариации физического типа человека во времени и пространстве» Я. Я. Рогинский и М. Г. Левин подчеркивают, что в советской науке принято строгое разделение этнографии и архео�логии, являющихся отраслями истории, и антропологии как области биологии. «Антропология есть отрасль естествознания, — пишут эти авторы, — которая изу�чает происхождение и эволюцию физической организации человека и его рас. Задача антропологии — проследить процесс перехода от биологических закономерностей, которым подчинялось существование животного предка человека, к закономерностям социальным...».
В научно-популярной и учебной литературе ход эволюции человека изображается обычно в виде чётких и легко запоминающихся схем, в которых процесс смены одних видов наших предков другими предстаёт, как простая линейная последовательность. Благодаря этому, всем известно, что сначала были австралопитеки, потом пителокантропы, их сменили неандертальцы, а они в последствии уступили место людям современного физического типа- Homo sapiens. Получается, что в своём развитии как будто просто поднимался вверх по лестнице, переходя в положенное время с одной ступеньке на другую и следуя при этом единственно возможным путём.
Этот процесс, действительно, развивался по восходящей, и на каждой его новой стадии появлялись существа, в чем-то превосходившие своих предшественников; Однако, при всей своей закономерности, тот вариант антропогенеза, который осуществился в реальности, все же не был ни единственно возможным, ни фатально неизбежным. Наша состоявшаяся эволюционная история — это только один из многих потенциально существовавших ее сценариев, который, в отличие от других, не остался запасным лишь в силу во многом случайного стечения мало связанных между собой обстоятельств.
Лишь сейчас, когда с непрерывно растущей горы фактов, открывается все более широкая панорама человеческого прошлого, начинаем мы понимать, что наша эволюционная история — это движение в лабиринте, что пройденный путь отнюдь не был единственно возможным и, наконец, что выбор того или иного поворота на этом пути часто определялся лишь случайным стечением случайных же обстоятельств.
Цель контрольной работы: изучить основные антропологические и социологические характеристики эпохи праобщины.
Задача: 1)охарактеризовать антропологические особенности Homo sapiens эпохи праобщины;
2)охарактеризовать социологические особенности Homo sapiens эпохи праобщины;
3)изучить модели, описывающие эволюцию человека.
Homo sapiens (человек разумный) - является научным названием для человеческого вида. Люди из семейства гоминид в отряде приматов, единственные живущее в наше время. От современных человекообразных, по мимо ряда анатомических особенностей, отличается значительной степенью развития материальной и нематериальной культуры (включая изготовление и использование орудий труда), способностью к членораздельной речи и развитому абстрактному мышлению.
Род Homo
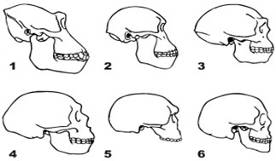
Рисунок 1 Вид черепа сбоку: 1. Горилла. 2. Австралопитек. 3. Homo erectus. 4. Неандерталец (Ля-Шапелль-о-Сен). 5. Штейнгеймский человек 6. Современный человек [5].
Слово homo в переводе с латыни означает «человек» и было выбрано для классификации ещё Линеем.
В латинском языке оно произошло от общеиндоевропейского корня «dhghem», земля.
В современной таксономии Homo sapiens — единственный ныне существующий вид рода Homo, хотя продолжающиеся исследования происхождения Homo sapiens дают всё новую и новую информацию о других видах Homo, давно уже вымерших. Некоторые из этих видов могли быть предками современных людей, но многие являются лишь «родственниками» и эволюционировали в сторону от нашего вида. В то же время продолжаются и дискуссии о том, какие из них считать отдельными видами, а какие — лишь расами одного вида. В некоторых случаях причиной разногласий является ограниченность или полное отсутствие необходимой информации, в других — различия в подходах к классификации.
Считается, что есть две главные научные школы, по-разному объясняющие движущие силы эволюции человека. Раньше других появились представления об адаптации предков людей, обитавших на деревьях, к жизни в саванне, куда они вышли для охоты на травоядных. Теория саванны, впервые сформулированная Раймондом Дартом, не исключает, что к саванне смогли приспособиться лишь те обезьяны, которые ещё в лесах приобрели необходимую для этого анатомию и поведенческие навыки.
Альтернативная гипотеза допускает, что человек эволюционировал под влиянием приспособления к земноводному существованию, то есть к собиранию моллюсков и прочей пищи на мелководье, что требовало, в частности, способности плавать и нырять, отличающей человека от прочих обезьян. Эта гипотеза объясняет многие анатомические особенности современного человека, такие как прямохождение, отсутствие шерсти, развитый слой подкожного жира, низкое положение гортани относительно носоглотки, характерное для морских млекопитающих, vernix caseosa или первородная смазка новорождённых детей, также характерная для морских млекопитающих, а не обезьян, крупный мозг, высокий нос, предотвращающий попадание воды в носоглотку и жирная кожа с обилием сальных желёз, которая может служить для защиты от воды.
Обсуждается несколько вариантов приспособления протолюдей к жизни в водной стихии, в том числе собирательство на мелководье и развитие новых способов передвижения в воде и доставки собранной пищи на берег, плавание и ныряние. Получить палеоантропологические доказательства земноводного обитания протолюдей крайне сложно, по крайней мере, из-за повышения уровня моря по окончании ледникового периода, из-за чего бывшее мелководье оказалось теперь на глубине 100-120 м. Однако археология и палеонтология позволяют исследовать рацион питания различных видов Homo и его влияние на эволюцию анатомии и поведения.
Появление Homo sapiens
Древнейшие представители вида Homo sapiens появились в результате эволюции 400—250 тыс. лет назад. Господствующей в наши дни гипотезой происхождения людей является африканская, согласно которой наш вид появился в Африке и оттуда распространился по всему свету, замещая существовавшие популяции H. erectus и H. neanderthalensis. Альтернативная гипотеза называется мультирегиональной. Согласно последней, люди, начиная, по-видимому, от H. erectus, эволюционировали как единый вид, в котором генные потоки могли свободно циркулировать. Имеющиеся в настоящее время данные палеоантропологии не позволяют сделать окончательный выбор между этими гипотезами, хотя данные генетики поддерживают африканскую теорию.
Сравнение полиморфизмов митохондриальной ДНК и датирование окаменелостей позволяют заключить, что Homo sapiens происходит из Африки, где около 200 тыс. лет назад жил последний общий предок ныне живущих людей по женской линии.
В 2009 году группа учёных под руководством Сары Тишкофф из Университета Пенсильвании опубликовала в журнале Science результаты комплексного исследования генетического разнообразия народов Африки. Они установили, что самой древней ветвью, испытавшей наименьшее количество смешиваний, как раньше и предполагалось, является генетический кластер, к которому принадлежат бушмены и другие народы, говорящие на койсанских языках. Скорее всего, они и являются той ветвью, которая ближе всего к общим предкам всего современного человечества.
Можно предположить, что 60 000 - 40 000 лет назад люди мигрировали в Азию, и оттуда в Европу (40 000 лет), Австралию и Америку (35 000 — 15 000 лет).
Антропогенез
Антропогенез — часть биологической эволюции, которая привела к появлению человека разумного (лат. Homo sapiens), отделившегося от прочих гоминид, человекообразных обезьян и плацентарных млекопитающих, процесс историко-эволюционного формирования физического типа человека, первоначального развития его трудовой деятельности, речи.
В эволюционном контексте термин «человек» относится не только к ныне живущим людям, но и к представителям вымерших видов рода Homo. Кроме того, исследования антропогенеза распространяются на других гоминид, например, австралопитеков. Род Homo отделился от австралопитеков или подобных им гоминид около 2 млн. лет назад в Африке. Существовало несколько видов людей, большинство из которых вымерло. К ним, в частности, относятся эректусы и неандертальцы.
Приматы — одна из старейших групп современных плацентарных млекопитающих. Эволюционная история приматов может быть прослежена примерно на 90 млн. лет назад, когда приматообразные разделились на приматов и шерстокрылов. 85 млн. лет назад сухоносые обезьяны отделились от мокроносых. Около 80 млн. л.н. разошлись линии долгопятообразных и обезьянообразных, а лемурообразные отделились от лориобразных. Останки древнейших приматов известны лишь из палеоценовых и эоценовых отложений Северной Америки, Евразии и Африки (плезиадаписы, антрасимия, нотарктус, дарвиниус (Ида) и др). Только для пургаториуса предполагается верхнемеловое время. После глобального похолодания, когда около 30 млн лет назад, в раннем олигоцене, Антарктида начала покрываться льдом, приматы вымерли повсеместно кроме Африки, Америки и юга Азии. Одним из выживших был грифопитек — ископаемая обезьяна, жившая на территории современной Германии и Турции около 16,5 млн. лет назад, на 1,5 млн. лет раньше, чем подобные виды появились в Африке. Возможно, первые человекообразные обезьяны также появились не в Африке, а в Евразии. С другой стороны, высказываются предположения, что предки гоминид мигрировали в Евразию из Африки около 17 млн лет назад, когда эти континенты некоторое время были соединены, прежде чем вновь разделились при расширении Средиземного моря. В начале миоцена (23.03 мл лет назад) климат снова стал тёплым, и они могли процветать в Евразии, после чего ареал одного из них, дриопитека, распространился из Европы или западной части Азии в Африку.
Возможно, что уцелевшая после климатических изменений тропическая популяция обезьян, хорошо представленная в слоях верхнего эоцена и нижнего олигоцена Файюмского оазиса в Египте (биретия, каранисия, сахарогалаго, протеопитек, египтопитек, катопитек и др.), дала начало всем ныне существующим приматам — лемурам Мадагаскара, лори Юго-Восточной Азии, африканским галаго, широконосым обезьянам Нового Света и узконосым обезьянам Старого Света (человекообразные обезьяны и мартышкообразные). В раннем миоцене примитивные узконосые обезьяны Восточной Африки пережили длительный период видообразования. Среди выделяемых ныне видов и родов обезьян этой эпохи известны, в частности, камойяпитек, моротопитек, лимнопитек, проконсул, афропитек, кениапитек, чорорапитек, экваториус, отавипитек, ньянцапитек, викториапитек, дендропитек, все из Восточной Африки, а также ореопитек, который обитал в Италии около 9 млн. лет назад и европейские — пиеролапитек, анойяпитек, дриопитек.
При сравнении ДНК современных обезьян было показано, что гиббоны отделились от общего ствола гоминид 18 млн. лет назад, а орангутаны — около 14 млн. лет назад. За исключением дендропитека останки ископаемых гиббонов науке неизвестны, и их происхождение остается неясным. Ископаемым прото-орангутаном считается сивапитек, обитавший в Азии около 12—10 млн. лет назад и хорапитек из Таиланда.
Предполагается, что видами, близкими к общему предку горилл, шимпанзе и людей, были накалипитек из Кении и уранопитек с Балканского полуострова. По данным молекулярной биологии, около 7 — 8 млн. лет назад сначала гориллы, а потом и шимпанзе отделились от предков людей. ДНК шимпанзе на 98,4 % идентична человеческой. Из-за влажного климата тропических лесов, в кислых почвах которого кости плохо сохраняются, а также отчасти из-за невнимания исследователей, сосредоточенных преимущественно на поиске предков человека, ископаемых горилл и шимпанзе до сих пор практически не найдено.
В середине миоцена климат снова стал холоднее и засушливее, что спровоцировало новое массовое вымирание фауны. Однако гоминины, как и многие другие виды (антилопы, гиены, собаки, свиньи, слоны, лошади) пережили климатические изменения и сумели к ним адаптироваться. Их дальнейшая эволюция привела к образованию множества новых видов.
Антропологические характеристики Homo sapiens эпохи праобщины
Черепная коробка.
У приматов и, особенно у гоминид размеры мозговой части черепа соответствуют объему черепной полости и определяются, преимущественно, величиной головного мозга. Несмотря на то, что что головной мозг заключен в твердую мозговую оболочку, взвешен в цереброспинальной жидкости и не соприкасается со стенками черепной полости, тем не менее рост покровных костей черепа, возникающих в первоначальной перепончатой оболочке, происходит в соответствии с разрастанием определенных участков поверхности коры головного мозга.
Увеличение полости черепа осуществляется в ходе эволюции гоминид двумя путями: перестройкой хрящевого основания черепа и разрастанием покровных костей перепончатого свода. У гоминид, перешедших к прямохождению, дорзальный отрезок основной кости передвигается задним концом вниз, образуя скат Блюменбаха, формируется задняя черепная ямка; затылочное отверстие из положения на задней поверхности черепа передвигается на нижнюю, и, по мере развития выступающего затылка, смещается вперед, приближая отвес из центра тяжести головы к линии опоры между затылочными мыщелками. Так как межмыщелковая линия и базион сохраняется не на всех ископаемых черепах, для ориентировки могут служить ушные отверстия, располагающиеся на несколько миллиметров впереди базиона. Проекция точки порион на линию назион – инион находится у сапиентных форм приблизительно на 50% расстояния от назиона, у среднеплейстоценовых – немного ближе к затылочному полюсу.
Мозговая коробка ранних гоминид сохраняет сходство с формой этого органа у наземных позвоночных – расширенной концевой полостью позвоночного канала. Удлиненная неширокая, уплощенная форма мозгового черепа у питекантроповых не сильно отличается от его конфигурации у наземных церкапитековых приматов, например, павианов сфинкс, хотя объем мозговой полости у питекантропа в 5,5 раза больше, чем у павиана.
Разрастание теменной доли мозга сопровождается у гоминид удлинением теменной кости и перемещением ее переднего конца - брегмы – во фронтальном направлении. У сапиентных гоминид отвес из брегмы находится немного впереди отвеса из пориона; у досапиентных форм брегма нередко сдвинута на несколько миллиметров назад.
Передняя (назальная) часть лобной кости образует расхождение наружной и внутренней пластинок кости, заключающее лобную пазуху. Прецеребральный отдел лобной кости в равной мере заполняется ячейками слизистой оболочки, проникающими из решетчатой кости.
Лобные полости у разных видов млекопитающих неоднородны как по расположению и по размерам, так и по содержанию костных пластинок и ячеек слизистой оболочки. В отряде приматов у некоторых родов церкопитековых и антропоморфных развитие пазух сопровождается образованием костного выступа передней стенки.
Общие размеры лицевого скелета у позднеплейстоценовых гоминид меньше, чем у питекантроповых. Однако подобное уменьшение охватывало не все группы. Так, для шапелльских гоминид характерны крупные челюсти; одновременно с ними, а, возможно, даже раньше, существовали ископаемые со средними размерами челюстей.
Кости скелета.
Известны лишь единичные находки, содержащие более или менее полный набор костей внечерепного скелета одного индивидуума.
В Триниле в 1891 г. Вместе с черепной крышкой питекантропа, но отдельно от нее была найдена бедренная кость, ничем не отличавшаяся от современной человеческой и имевшая к тому патологическое разрастание верхней части диафиза. Основываясь на этой находке, Дюбуа наименовал открытое им ископаемое «питекантроп прямоходящий».
Если эопрейстоценовые гоминиды освоили прямохождение в основных его элементах, то тринильская находка устанавливает, что в нижнем плейстоцене существовали гоминиды с более полным комплексом приспособлений к двуногой локомоции и, в первую очередь, с бедренной костью, приспособленной к ортоградности.
Бедренная кость синатропов также не имеет видоразграничительных особенностей структуры, но отличается от современного типа большей массивностью.
До открытия прегоминид австралопитековай группы все особенности строения скелета ископаемых гоминид рассматривались как показатели близости ископаемого к группе шимпанзе – горилла, в которой видели наиболее сохранившийся вариант общего исходного типа. На данный момент, следует признать, что многоие особенности скелета гоминид имеют сравнительно позднее происхождение, их формирование шло в процессе развития структурных корреляци, сопровождавших прямохождение. Что касается исходной формы гоминид, то она сохраняла при переходе к прямохождению некоторые особенности прототипа высших приматов, в частности в стопе и кисти некоторое отведение первого луча и меньшую его массивность. У нижне и среднеплейстоценовых гоминид, а иногда и у позднеплейстоценовых, можно установить отдельные признаки незаконченной выработки системы корреляций (в развитии мыщелков, большой берцовой кости, в рельефе плечевой кости, в строении запястья и предплюсны), но эти признаки выражены не настолько единообразно, что бы стать отличительной чертой той или иной группы ископаемых гоминид плейстоцена.
Речь и мышление.
В обыденной жизни речью, или языком, называют самые разнообразные звуки, производимые голосовыми органами, а показателем мышления считают всякий целесообразный акт деятельности. В научном исследовании единицу речевой деятельности составляет «слово», а единицу мышления – «понятие» разной сложности. Понятие и слово находятся в неразрывной связи. Звуки, не выражающие понятие, нельзя назвать словами, а понятия не могут возникать без движений голосовых органов, независимо от того, произносятся ли слова вслух или мысленно.
Начальная стадия формирования слов включала многие элементы структуры голосовых органов и головного мозга. Наиболее существенными этапами преобразования звукового аппарата животных в человеческие органы речи следует считать загиб корня языка в гортанную полость, усиление голосовых связок, разрастание внутренних краев черпаловидных хрящей и ряд других преобразований, приведших к более четкой дифференциации издаваемых звуков, разделение выдыхаемого воздуха на верхнюю, или носовую, струю и нижнюю, или ротовую. Увеличение подвижности языка открывало возможность разнообразных изгибов его тела и образование преград выдыхаемому воздуху, то есть дифференциации гортанных, небных, губных и других звуков.
Существенное значение в развитии речевой способности имело уменьшение размеров нижней челюсти и ее мускулов: при быстрой смене артикуляции массивная нижняя челюсть и мускулатура создавали бы большую инерцию в работе речевого аппарата, основанной, как известно, именно на быстрой смене артикуляции. Надо полагать, что арханотропы вследствие массивности челюстей могли произносить только небольшое число различных выкриков.
Дальнейший ход развития состоял в продолжающейся редукции нижнечелюстного аппарата, а также в дальнейшем утоньшении голосовых органов, выделении особого голосового мускула и появлении многих других структурных особенностей. Из многих сотен звуков в ходе эволюции речевого аппарата постепенно выработалось около трех десятков, получавших смысловую нагрузку, то есть связанных с определенными представлениями и понятиями. Существенная черта в эволюции речи – способность образовывать и быстро заменять разные сочетания отдельных звуков.
На одном из последних этапов развития речевой и мыслительной функции происходило усиленное разрастание прецентральной, нижнелобной, верхневисочной и верхнелобной извилин, а затем и цитоархитектонических полей, координирующих различные возбуждения и связывающих их с возбуждениями голосовых органов.
Промежуточную стадию между выкриками и хотя бы начальными словами называют лалиями (лепетом).
Сравнительное изучение эндокранов ископаемых гоминид указывает на сравнительно малое развитие цитоархитектонических полей, связанных с речевыми функциями. У ископаемых, относимых к группе палеоантропов, эндокраны несут ясные отличия от современных.
Простейшие формы связи – соединение в одном речении субъекта и предиката. Возникновение синтагмов (связи между элементами мысли) следует определить как один из кардинальных этапов эволюции речи.
Следует полагать, что люди верхнего палеолита уже овладели синтагматической формой речи. Связь нескольких синтагмов, образование суждений, дифференциация частей речи и ее грамматических элементов получила развитие в конце палеолита или в последующие периоды.
Социологические характеристики Homo sapiens эпохи праобщины
Развитие коллективизма.
Выделение человека из животного мира стало возможным только благодаря труду, который сам по себе представлял коллективную форму воздействия человека на природу.
На самых ранних этапах антропогенеза и истории первобытного общества имело место регулирование в добывании и распределении пищи, половой жизни и так далее.
Развитие загонной охоты, совместная защита от хищных животных, поддержание огня – все это способствовало консолидации праобщины, развитию начала инстинктивных, а затем и осознанных форм взаимопомощи. В этом же направлении сплочения коллектива действовало и усовершенствование языка. Особенно большой прогресс приходится на заключительный этап существования праобщины – мустьерское время. Именно к этому времени относят первые яркие свидетельства заботы о членах коллектива – неандертальские погребения.
Охота в развитии праобщины.
Охота была более прогрессивной отраслью хозяйства, во многом определившей развитие первобытных человеческих коллективов.
Объектами охоты в зависимости от фауны той или другой области были различные животные. В тропической зоне это были гиппопотамы, тапиры, антилопы, дикие быки и так далее. Иногда среди костей животных, обнаруженных на ашельских стоянках, попадаются кости даже слонов. В более сереных районах охотились на лошадей, оленей, кабанов, зубров, иногда убивали и хищников – пещерных медведей и львов, мясо которых также шло в пищу. В высокогорной зоне преимущественную роль в охоте, например у неандертальцев, играла добыча горных козлов, что видно из находок в пещере Тешик-Таш. О размерах охоты в какой-то степени можно судить на основе подсчета костей. Культурный слой многих из них содержит остатки сотен, а иногда даже тысяч животных.
Охоту на крупных животных трудно представить себе без загонного способа. Вооружение ашельского охотника было слишком слабым, чтобы он мог убить крупное животное непосредственно. Вероятно животных пугали шумом, огнем, камнями и, как показывает местоположение многих стоянок, гнали к глубокому ущелью или большому обрыву. Животные падали и разбивались, и человеку оставалось только добить их. Вот почему именно охота, и прежде всего охота на крупных животных, была той формой трудовой деятельности, которая больше всего стимулировала организованность праобщины, заставляла ее членов все тесне сплачиваться в трудовом процессе и демонстрировала им силу коллективизма.
Охота была наиболее эффективным источником получения мясной пищи. Мясо, содержащее важнейшие для человеческого организма вещества – белки, жиры и углеводы, не только было сытной пищей, особенно после обработки его на огне, но ускоряло рост и повышало жизнедеятельность первобытного человека. Это обстоятельство было подчеркнуто уже Энгельсом, видевшим в мясной пище важнейший стимул биологического прогресса на ранних ступенях человеческой эволюции.
Для добывания пищи древние гоминиды нуждались в разных орудиях – ударных, метательных, рычаговых, режущих и скоблящих. Доступный материал для различных поделок доставляли необходимые в природе сучки, палки, раковины моллюсков, кости крупных млекопитающих, камни. В зависимости от места обитания наиболее доступными оказывались разные материалы, но едва ли древний человек мог обойтись без древесных сучков, чтобы захватить или придавить мелкое животное, без палок разной длинны, служивших для сбивания плодов, для раскопки норы и клубней. Для раскопки и для проколов шкуры убитого животного могли служить рога с острыми концами. Длинные кости животных, расколотые поперек диафиза, могли быть использованы для отделения шкуры от туши, для перерезки сухожилий. Однако для срезания и скобления более пригодны были камни с заостренными гранями. Камни служили для удара и для метания при нападении или защите.
Древний гоминид использовал для изготовления орудий все доступные материалы и при том не только в начальную эоплейстоценовую эпоху, но и во все последующие. Дерево и кость в среднем и позднем палеолите служили для изготовления отщепов камня, пластинчатых орудий. Деревянные копья в самом примитивном виде принадлежат к древнейшему вооружению человека, ставшему особо эффективным в ашельское время, когда концы древка можно было заострять обжиганием. Копья со вставленными в древко острыми обломками кости или камня были действенным оружием на протяжении тысячелетий.
Половые отношения в праобщине.
Одной из основных линий борьбы биологических и социальных начал в праобщине были отношения по детопроизводству, или половые отношения.
Часть ученых считает, что праобщина как начальная форма общественной организации могла возникнуть лишь в результате растворения в ней зоологических семей и взаимной терпимости взрослых самцов, то есть установления нерегламентированных неупорядоченных половых отношений.
Однако существует и другая точка зрения, по которой праобщина унаследовала от предшесвовавших ей животных объединений гаремную семью со свойственной ей регламентацией половой жизни.
Культура первобытного общества.
Сначала в стаде первобытных людей, а затем с формированием Homo sapiens на протяжении всего развития родового стоя в процессе трудовой деятельности человека шло накопление полезных знаний, вырабатывались формы духовной культуры. Огромную роль в накоплении полезных знаний об окружающей природе сыграли собирательство и охота. Эти знания передавались из поколения в поколение.
В первобытном обществе были разработаны первые формы связи между людьми, находящимися на далеком расстоянии друг от друга. Наиболее простой и древней формой связи являлась световая сигнализация. Была также создана система условных знаков, вырезавшихся на коре деревьев или составлявшихся из веток или камней, с помощью которых передавалась информация охотникам. Использовались шнуры с узлами, имевшим условное значение. Затем у ряда народов появилась самая ранняя форма письменности – пиктография, или рисуночное письмо.
Концепции моноцентизма и полицентризма
Существует мнение, что эволюцию человека описывают только две модели: моноцентризм и полицентизм. На самом деле эти модели очень старые – им около ста лет. Они перестали использоваться антропологами примерно полвека назад.
Моноцентризм утверждает, что человечество произошло из единого центра (например, Восточная Африка, Индостан или Сунда, а может и вовсе Атлантида). Полицентризм убеждает, что у каждой расы был свой путь эволюции от ранних гоминид, а возможно и от древесных приматов, и между этими стволами нет никакой генетической связи.
Согласно теории полицентризма, современные расы человека возникли в результате длительной параллельной эволюции нескольких филетических линий на разных материках: европеоидная в Европе, негроидная в Африке, монголоидная в Центральной и Восточной Азии, австралоидная в Австралии. Однако если эволюция расовых комплексов и шла параллельно на разных континентах, она не могла быть полностью независимой, поскольку древние проторасы должны были скрещиваться на границах своих ареалов и обмениваться генетической информацией. В ряде областей сформировались промежуточные малые расы, характеризующиеся смешением признаков разных больших рас. Так, промежуточное положение между европеоидной и монголоидной расами занимают южносибирская и уральская малые расы, между европеоидной и негроидной — эфиопская и т. д.
С позиций моноцентризма современные человеческие расы сформировались относительно поздно, 25—35 тыс. лет назад, в процессе расселения неоантропов из области их возникновения. При этом также допускается возможность скрещивания (хотя бы ограниченного) неоантропов во время их экспансии с вытесняемыми популяциями палеоантропов (как процесса интрогрессивной межвидовой гибридизации) с проникновением аллелей последних в генофонды популяций неоантропов. Это также могло способствовать расовой дифференциации и устойчивости некоторых фенотипических признаков (подобных лопатообразным резцам монголоидов) в центрах расообразования.
Существуют и компромиссные между моно- и полицентризмом концепции, допускающие расхождение филетических линий, ведущих к разным большим расам, на различных уровнях (стадиях) антропогенеза: например, более близких друг к другу европеоидов и негроидов уже на стадии неоантропов с первоначальным развитием их предкового ствола в западной части Старого Света, тогда как еще на стадии палеоантропов могла обособиться восточная ветвь — монголоиды и, может быть, австралоиды.
Большие расы человека занимают обширные территории, охватывающие народы, которые различаются по уровню экономического развития, культуре, языку. Нет сколько-нибудь четких совпадений понятий «раса» и «этнос» (народ, нация, национальность). В то же время есть примеры антропологических типов (малые, а иногда и большие расы), которые соответствуют одному или нескольким близким этносам, например, лапаноидная раса и саамы. Гораздо чаще, однако, наблюдается обратное: один антропологический тип широко распространен среди многих этносов, как, например, в коренном населении Америки или у народов Северной Европы. Вообще все крупные народы, как правило, неоднородны в антропологическом отношении. Нет также совпадения между расами и языковыми группами — последние возникали позже, чем расы. Так, среди тюркоязычных народов имеются представители как европеоидов (азербайджанцы), так и монголоидов (якуты). Термин «расы» неприменим к языковым семьям — например, нужно говорить не о «славянской расе», а о группе родственных народов, говорящих на славянских языках.
Заключение
В последние десятилетия почти каждый год приносит антропологические открытия — ископаемые останки все новых приматов, в том числе и гоминид. Возникают споры об их положении на эволюционном древе и приоритете в отношении родства с современным человеком. Эти открытия революционизируют науку и вызывают желание пересмотреть все старые представления, вплоть до учения Ч.Дарвина и концепции Ф.Энгельса. Между тем, эти открытия скорее указывают на огромное разнообразие семейств и видов в отряде приматов, чем проясняют преемственность форм в схеме их эволюции. В учении о человеке до сих не выработан единый методологический подход, хотя необходимость его осознана достаточно давно, и мы находимся буквально у порога решения этой проблемы. По-видимому, решена она будет тогда, когда сложится тенденция специализации по проблемам, а не по наукам.
Таким образом, мы, изучив основные антропологические и социологические характеристики эпохи праобщины, пришли к выводу, что, чем больше наука узнает о человеке, тем более серьезные задачи возникают перед ней, в частности, все более актуальными становятся проблемы изучения механизмов работы мозга, специфики эстетического чувства, семиотических оснований культуры и др.
Список литературы
1. Алексеев В.П., Першиц А.И. История первобытного общества. М., 1990.
2. Анисимов А.Ф. Исторические особенности первобытного мышления. Л., 1971.
3. Бунак В.В. Род Homo, его возникновение и последующая эволюция. М., 1979.
4. Горемыкина В.И. История первобытного общества. Минск, 1973.
5. Семенов Ю.И. На заре человеческой истории. М., 1989.
|
 Скачать 58.43 Kb.
Скачать 58.43 Kb.