Открытие структуры ДНК. Классические эксперименты днк как генетический материал
 Скачать 5.29 Mb. Скачать 5.29 Mb.
|

|
https://ru.khanacademy.org/science/biology/dna-as-the-genetic-material/dna-discovery-and-structure/a/classic-experiments-dna-as-the-genetic-material Классические эксперименты: ДНК как генетический материалЭксперименты Фредерика Гриффита, Освальда Эвери и его коллег, а также Альфреда Херши и Марты Чейз. ВведениеНаше современное представление о ДНК и наследственности привело к разработке большого количества направлений, таких как криминалистический анализ, тестирование на отцовство и генетический скрининг. Поэтому благодаря такому широкому применению, сегодня многие люди имеют базовые представление о том, что такое ДНК. Удивительно, но менее ста лет назад даже лучшие учёные не знали, что ДНК отвечает за наследственность! В этой статье мы рассмотрим несколько классических опытов, которые привели к пониманию того, что ДНК является носителем генетической информации. Протеины и ДНКСогласно работе Грегора Менделя признаки (такие как окраска лепестков у цветков гороха) не наследуются напрямую, а скорее определяются генами, передаваемыми от родителей к потомкам. Последующие работы ученых, среди которых Теодор Бовери, Уолтер Саттон и Томас Хант Морган, доказали, что наследственные признаки Менделя, скорее всего, связаны с хромосомами. Изначально ученые считали, что белки, которые содержатся в хромосомах вместе с ДНК, являются генетическим материалом. Ведь было известно, что белки имеют различные аминокислотные последовательности, в то время как ДНК считалась скучным, повторяющимся полимером, отчасти из-за неправильного (но распространенного) понимания его структуры и состава ^11start superscript, 1, end superscript. Сегодня нам известно, что изначальное представление о ДНК было ошибочным. В действительности ДНК несёт большой объём информации и мы расскажем об этом подробнее в статье открытие структуры ДНК. Но как учёные впервые поняли, что «скучная» ДНК на самом деле является генетическим материалом? Фредерик Гриффит: Трансформация бактерийВ 1928 году британский бактериолог Фредерик Гриффит провел серию экспериментов с использованием бактерий Streptococcus pneumoniae (пневмококк) и мышей. Гриффит не пытался определить генетический материал, а пытался разработать вакцину против пневмонии. В своих экспериментах Гриффит использовал два родственных штамма бактерий, известных как R и S. R-штамм. При выращивании в чашке Петри R-бактерии образовывали колонии или скопления родственных бактерий, которые имели четко очерченные края и шероховатый внешний вид (аббревиатура «R» от английского "rough" - грубый, шероховатый). R-бактерии были невирулентными, то есть не вызывали болезни при введении мышам. S-штамм. S-бактерии образовывали округлые и гладкие колонии (S — это первая буква английского слова «smooth» — «гладкий»). Их гладкость была обусловлена оболочкой из полисахарида, создаваемой бактериями. Эта оболочка защищала S-бактерии от иммунной системы мышей, делая их вирулентными (способными вызывать заболевания). У мышей, которым вводили живые S-бактерии, развивалась пневмония, и они умирали. В рамках своих экспериментов Гриффит попытался ввести мышам инактивированные при нагревании S-бактерии (то есть S-бактерии нагрели до очень высокой температуре, что привело к разрушению клетки). Неудивительно, что такие S-бактерии не вызывали заболевания у мышей. Однако эксперименты приняли неожиданный оборот, когда безвредные R-бактерии были введены мышам вместе с ранее нагретыми S-бактериями. Мало того, что у мышей развилась пневмония и они погибли, но когда Гриффит взял у мёртвой мыши образец крови, он обнаружил, что в нём содержатся живые бактерии S-штамма! 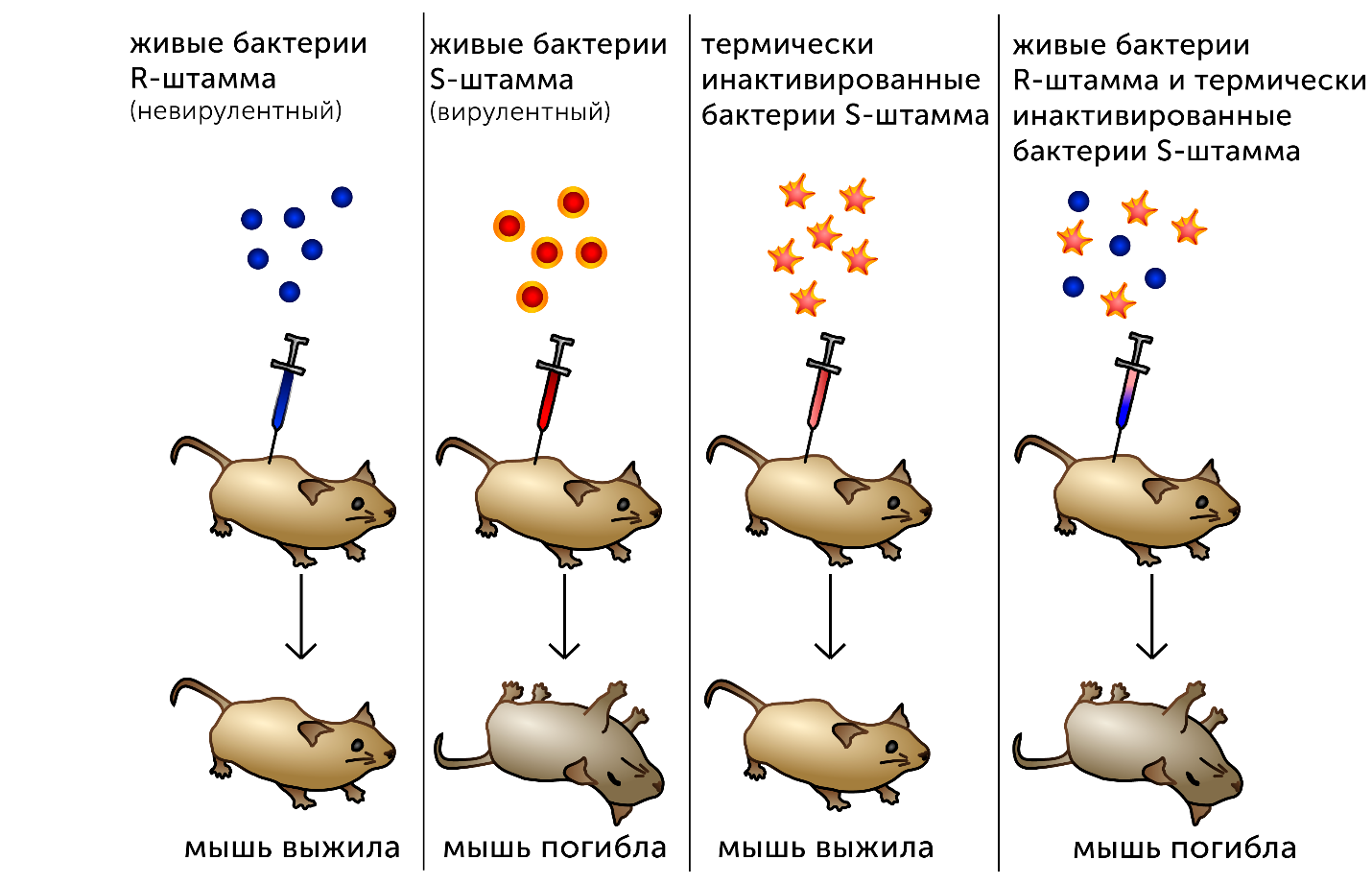 Таким образом, Гриффит пришел к выводу, что бактерии R-штамма, должно быть, переняли что-то от инактивированных S-бактерий, и это что-то он назвал «фактором трансформации». Благодаря этому R-бактерии смогли «трансформироваться» в бактерии с гладкой поверхностью и стать вирулентными. Эйвери, Маккарти и Маклауд: выявление фактора трансформацииВ 1944 году трое канадских и американских исследователей, Освальд Эвери, Маклин Маккарти и Колин Маклауд, решили разобраться с «фактором трансформации» Гриффита. Для этого они начали с объёмных культур S-клеток, и после долгой последовательности биохимических этапов (тщательно определённых экспериментальным путем) они постепенно выделили фактор трансформации через отмывания, разделения или ферментативного разрушения других клеточных компонентов. Благодаря этому методу они смогли получить небольшое количество высокоочищенного фактора трансформации и в результате ряда других анализов определить, что он из себя представляет ^22squared Эвери и его коллеги предложили несколько прямых доказательств того, что фактором трансформации может быть ДНК^22squared: Очищенное вещество в химических тестах на обнаружения белков дало отрицательный результат, но при этом также дало явный положительный результат в химических тестах на обнаружение ДНК. Состав очищенного фактора очень напоминал ДНК по соотношению азота и фосфора. Ферменты, разрушающие белки и РНК, на него практически не влияли, однако ферменты, способные расщеплять ДНК, устраняли трансформирующую активность. Получившиеся результаты указывали на то, что ДНК ,вероятно, является фактором трансформации. Однако, Эйвери был осторожен в интерпретации своих результатов. Он понимал, что самим фактором трансформации может быть какое-то дополнительное вещество, присутствующее в небольших количествах, а не ДНК^33cubed. И поскольку такая возможность оставалась, споры о роли ДНК продолжались до 1952 года, когда Альфред Херши и Марта Чейз, используя другой подход, смогли окончательно установить, что именно ДНК является носителем генетического материала. Эксперименты Херши-ЧейзВ своих экспериментах, ставших теперь легендарными, Херши и Чейз изучали бактериофаги — вирусы, поражающие бактерии. Используемые ими фаги были простыми частицами, состоящими из внешней белковой оболочки и заключённой внутри ДНК. Херши и Чейз знали, что фаги прикрепляются к поверхности бактериальной клетки-хозяина и вводят в неё некое вещество (ДНК или белок). Это вещество содержало «инструкции», которые заставляли бактерию производить много новых фагов, то есть в клетку попадал генетический материал фага. Однако до эксперимента Херши полагал, что генетический материал окажется белком^44start superscript, 4, end superscript. Чтобы установить, что именно фаг вводит в бактерию-хозяина, ДНК или белок, Херши и Чейз подготовили две разные партии фагов. В каждой партии фаг наращивался в присутствии определенного радиоактивного элемента, который был включён в состав макромолекул фага (ДНК и белков). Один образец фага был получен в присутствии ^{35}\text S35Sstart superscript, 35, end superscript, start text, S, end text (радиоактивного изотопа серы-35). Сера содержится во многих белках, но отсутствует в ДНК, поэтому только фаговые белки были радиоактивно мечены таким способом. Другой образец был получен в присутствии ^{32}\text P32Pstart superscript, 32, end superscript, start text, P, end text, (радиоактивного изотопа фосфора-32). Фосфор обнаружен в ДНК, но не в белках, поэтому только фаговая ДНК (а не фаговые белки) была радиоактивно мечена. Каждая партия фага была использована для заражения отдельных культур бактерий. После заражения каждую культуру помещали в центрифугу и вращали на большой скорости, чтобы разделить бактерии и фаги. Центрифугирование заставляет более тяжёлый материал, такой как бактерии, опускаться на дно пробирки и выпадать в виде осадка. Более лёгкий материал, например, среда, используемая для выращивания культур, наряду с фагами, остается на поверхности и образует жидкий слой, называемый супернатантом 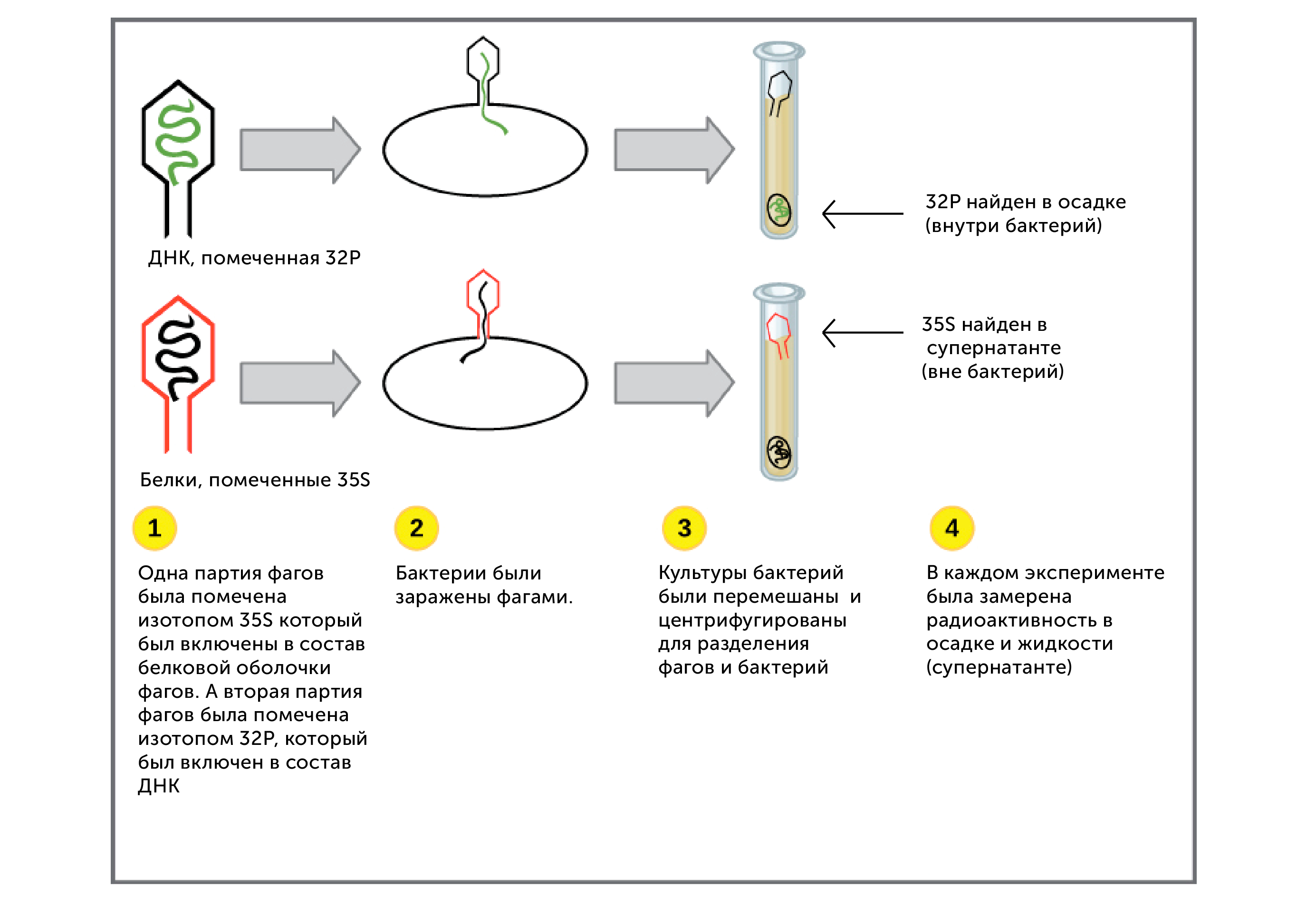 Одна партия фагов была отмечена 35S(изотоп серы-35), который встраивался в белковую оболочку. Другая партия была отмечена 32P(изотоп фосфор-32), который встраивался в ДНК. Бактерии были заражены фагами. Культуры перемешивали и центрифугировали для разделения фагов от бактерий. Радиоактивность измеряли в осадке и жидкости (супернатант). 32P (изотоп фосфор-32) был обнаружен в осадке (внутри бактерий), в то время как 35S (изотоп серы-32) был обнаружен в супернатанте (вне бактерий) _изображение модифицировано, источник: "Исторический контекст современного понимания: Изображение 3," by OpenStax College, Biology (CC BY 3,0)._ Когда Херши и Чейз измерили радиоактивность в осадке и супернатанте в обоих экспериментах, они обнаружили, что в осадке оказалось большое количество изотопа ^{32}\text P32Pstart superscript, 32, end superscript, start text, P, end text, тогда как почти весь изотоп ^{35}\text S35Sstart superscript, 35, end superscript, start text, S, end text находился в супернатанте. Основываясь на этом и других экспериментах, Херши и Чейз пришли к выводу, что в клетки была введена именно ДНК, а не белок, и именно она являлась генетическим материалом фага. [Верно ли это для всех вирусов?] Оставшиеся вопросыРаботы исследователей, приведенные выше, предоставили убедительные доказательства того, что ДНК является генетическим материалом. Однако результаты этих работы оставили без ответа вопрос: как такая, казалось бы, простая молекула может кодировать генетическую информацию, необходимую для построения сложного организма. Дальнейшие исследования многих ученых, в том числе Эрвина Чаргаффа, Джеймса Уотсона, Фрэнсиса Крика и Розалинд Франклин, привели к открытию структуры ДНК, и пониманию того, как ДНК может кодировать большие объемы информации. Открытие структуры ДНК Структура двойной спирали ДНК и как она была открыта. Чаргафф, Уотсон и Крик, Уилкинс и Франклин. https://ru.khanacademy.org/science/biology/dna-as-the-genetic-material/dna-discovery-and-structure/a/discovery-of-the-structure-of-dna ВведениеСегодня двойная спираль ДНК является, пожалуй, самой известной биологической молекулой. Строятся похожие на неё лестницы, небоскрёбы, украшения и пешеходные мосты (на иллюстрации ниже показан такой мост в Сингапуре) и многое другое. Здесь сложно не согласится с архитекторами и дизайнерами: ведь двойная спираль - это действительно красивая структура, в которой сочетается форма и функциональность. Однако двойная спираль далеко не всегда была частью нашего культурного словаря. Фактически, до 1950-х годов структура ДНК оставалась загадкой.  Иллюстрация: оригинал взят из: "Мост Хеликс, Сингапур," by William Cho, CC BY-SA 2,0 В этой статье мы кратко рассмотрим, как Джеймс Уотсон, Фрэнсис Крик, Розалинд Франклин и другие учёные открыли двойную спиральную структуру ДНК, а затем поговорим о её свойствах. Компоненты ДНКБиохимик Фибус Левин и другие ученые-современники Уотсона и Крика к тому времени показали, что ДНК состоит из компонентов, называемых нуклеотидами^11start superscript, 1, end superscript. Каждый нуклеотид состоит из сахара (дезоксирибозы), фосфатной группы и одного из четырех азотистых оснований: аденина (A), тимина (T), гуанина (G) или цитозина (C). Основания C и T, имеющие в своей структуре только одно углеродное кольцо, называются пиримидинами, а основания A и G, имеющие два кольца, называются пуринами. 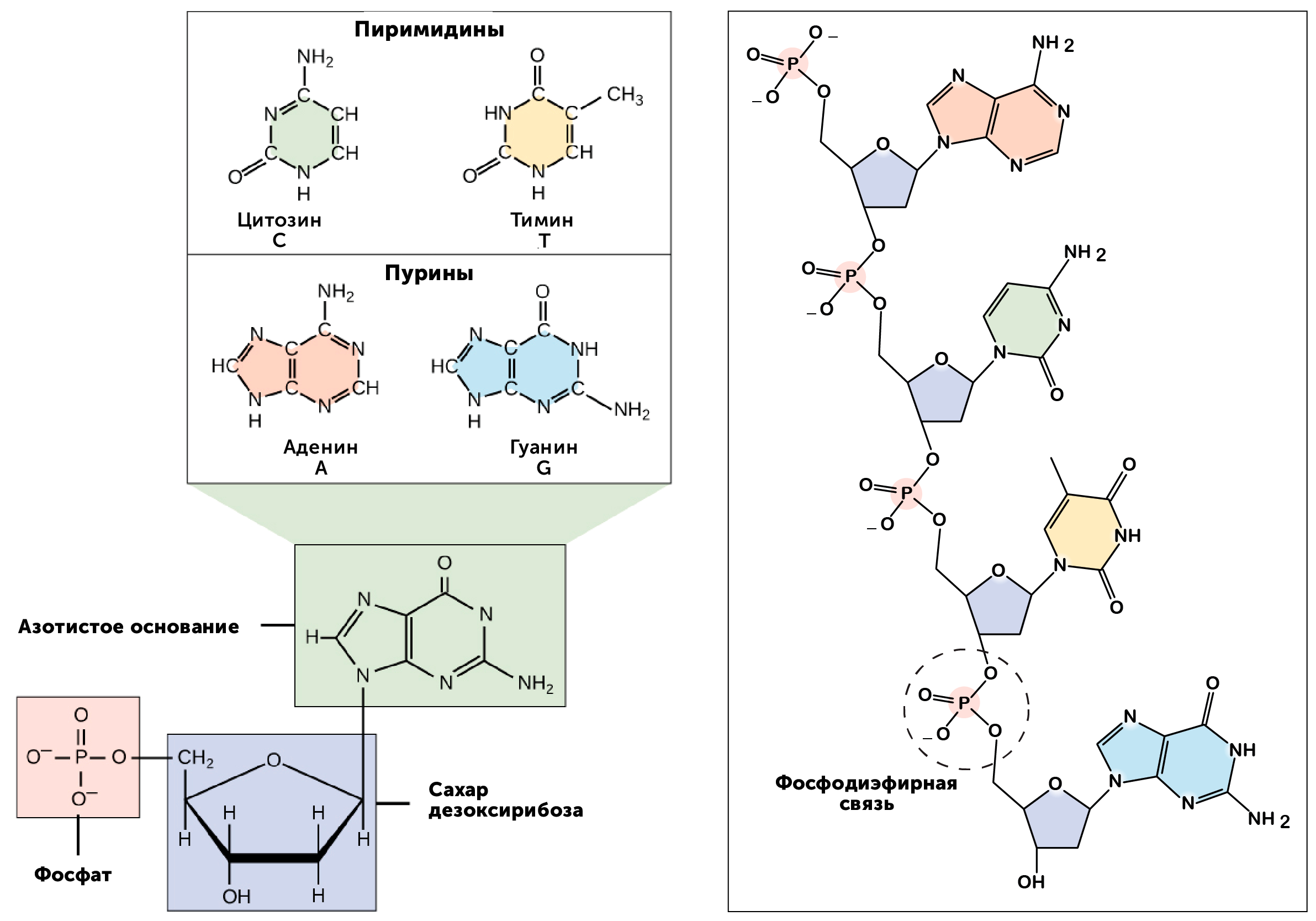 Слева: структура нуклеотида ДНК. Сахар – дезоксирибоза, соединен с фосфатной группой и азотистым основанием. Основание может быть любым из четырех возможных вариантов: цитозин (С), тимин (Т), аденин (А) и гуанин (G). Четыре основания различаются по структуре и функциональным группам. Цитозин и тимин являются пиримидинами и имеют только одно углеродное кольцо. Аденин и гуанин являются пуринами , это значит, что в их структуре есть два углеродных кольца. Справа: цепь соединенных нуклеотидов ДНК. Сахара в составе различных нуклеотидов связаны между собой фосфодиэфирными связями. Эта связь состоит из фосфатной группы, в которой два атома кислорода связаны с другими атомами - в данном случае с атомами углерода соседних дезоксирибозных сахаров. Нить ДНК состоит из чередующихся фосфатных групп и дезоксирибозных сахаров (сахарофосфатного остова), причем азотистые основания присоединены к дезоксирибозным сахарам. Иллюстрация: левая сторона, изображение изменино, источник: "Нуклеиновая кислота: Изображение 1," от OpenStax College, Biology (CC BY 3,0). Правая сторона, изображение изменино, источник: "Химическая структура ДНК," от Madeleine Price Ball (CC0/public domain). Нуклеотиды ДНК собираются в цепочки, связанные ковалентными связями, которые образуются между дезоксирибозным сахаром одного нуклеотида и фосфатной группой следующего. Такое расположение создаёт чередующуюся цепочку дезоксирибозных сахаров и фосфатных групп в полимере ДНК — так называемый сахарофосфатный остов Правила ЧаргаффаЕщё одну важную информацию о структуре ДНК узнал австрийский биохимик Эрвин Чаргафф. Он проанализировал ДНК нескольких видов и выяснил, что она состоит из оснований A, T, C и G.Также он сделал несколько важный наблюдений: Основания A, T, C и G не были найдены в равных количествах (как предсказывали некоторые модели того времени) Количество оснований варьировалось у разных видов, но не у особей одного и того же вида. Количество A всегда равнялось количеству T, а количество C всегда равнялось количеству G (A = T и G = C) Эти открытия, получившие название правила Чаргаффа, сыграли решающую роль в построении схемы двойной спирали ДНК Уотсоном и Криком. Уотсон, Крик и Розалинд ФранклинВ начале 1950-х годов американский биолог Джеймс Уотсон и британский физик Фрэнсис Крик опубликовали свою знаменитую модель двойной спирали ДНК. Они оказались первыми, кто пересёк «финишную прямую» в этой научной «гонке», в то время как другие учёные, например, Лайнус Полинг, открывший вторичную структуру белка, также пытались найти корректную модель. Вместо того, чтобы проводить новые эксперименты в лаборатории, Уотсон и Крик в основном собирали, анализировали и объединяли уже существующие данных^22squared. Некоторые из наиболее важных подсказок для структуры ДНК пришли от Розалинд Франклин, женщины биохимика, работавшей тогда в лаборатории физика Мориса Уилкинса. Франклин была экспертом в области определения структуры молекул, известного как рентгеновская кристаллография. Если кристаллизовать молекулу (например, ДНК) и облучить ренгтеновскими лучами, некоторые лучи преломляются атомами и образуется дифракционная картина, по которой можно судить о строении молекулы.  Рентгенограмма ДНК. Дифракционная картина имеет форму X, представляющую двухцепочечную спиральную структуру ДНК. Изображение изменено, источник: "Структура ДНК и секвенирование: Изображение 2," от OpenStax College, Biology (CC BY 3,0) Рентгенограммы Розалинд Франклин подсказали Уотсону и Крику, как в действительности устроена ДНК. Важную роль в этом открытии сыграла знаменитая «фотография 51» — удивительно чёткая рентгенограмма дифракционной картины ДНК, созданная Франклин и её аспирантом. (Современный пример дифракционной картины, создаваемой ДНК, показан выше.) Увидев Х-образную дифракционную картину на изображении, полученном Франклин, Уотсон сразу предположил, что молекула ДНК имеет спиральную двухцепочечную структуру^33cubed. [Уотсон и Крик украли данные Франклин?] Уотсон и Крик собрали вместе данные многих исследователей (в том числе Франклин, Уилкинса, Чаргаффа и других), и на основании этих данных построили свою знаменитую модель трехмерной структуры ДНК. В 1962 году Джеймс Уотсон, Фрэнсис Крик и Морис Уилкинс были удостоены Нобелевской премии по медицине. К сожалению, к тому времени Франклин умерла, а, как известно, Нобелевские премии не присуждаются посмертно. Модель ДНК Уотсона и КрикаСтруктура ДНК, в модели Уотсона и Крика, представляет собой двухцепочечную антипараллельную правостороннюю спираль. Сахарофосфатный остов цепей ДНК образует внешнюю часть спирали, в то время как азотистые основания находятся внутри и образуют пары, связанные водородными связями, которые удерживают две цепочки ДНК вместе. В приведенной ниже модели оранжевые и красные атомы обозначают фосфаты в сахарофосфатном остове, а синие атомы внутри спирали принадлежат азотистым основаниям. 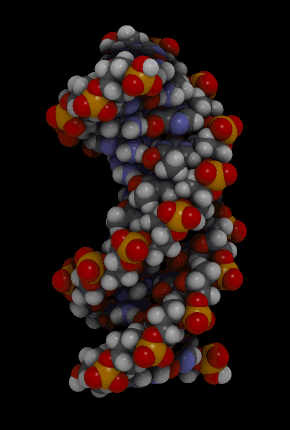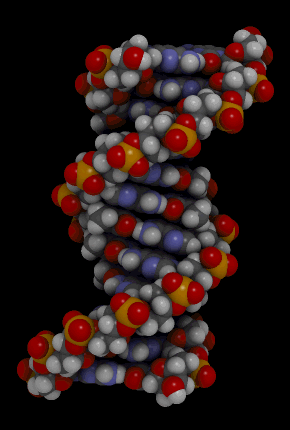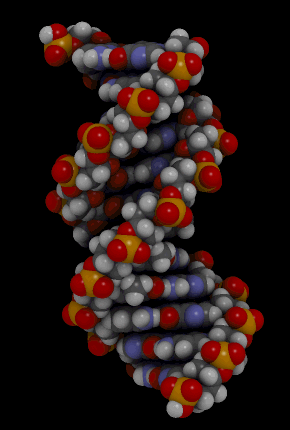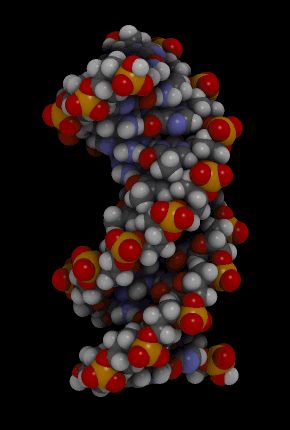 3D анимация молекулярной структуры двойной спирали ДНК. Источник изображения: "Bdna cropped," от Jahobr, public domain. Антипараллельная ориентация цепей ДНКДвухцепочечная ДНК представляет собой антипараллельную молекулу, что означает, что она состоит из двух нитей, которые находятся рядом друг с другом, однако направлены в противоположных стороны. В двухцепочечной молекуле ДНК 5'-конец (читается: пять-штрих конец)(фосфатсодержащий конец) одной нити совпадает с 3'-концом (гидроксилсодержащий конец) второй нити, и наоборот. [В чём смысл «штрихов» в обозначениях «3’» и «5‘»?] 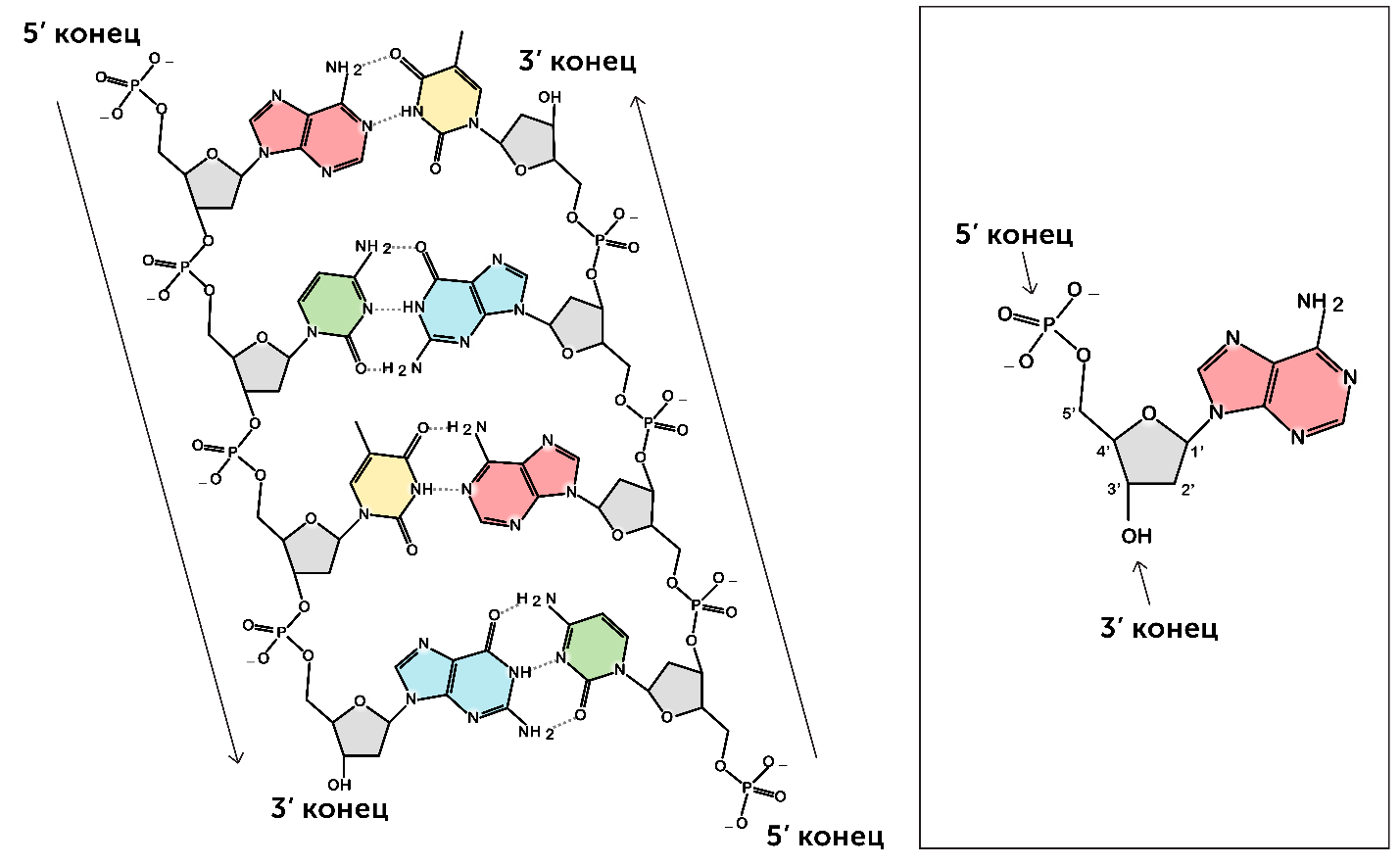 Слева: изображение антипараллельной структуры ДНК. Показан короткий сегмент двойной спирали ДНК, состоящий из двух нитей, удерживаемых вместе водородными связями между азотистыми основаниями. Нить слева имеет фосфатную группу, в верхней части (5'-конец), и гидроксильную группу, в нижней части (3 'конец). Нить справа имеет противоположную ориентацию, с фосфатной группой, внизу (5'-конец), и гидроксидом, наверху (3'-конец). Таким образом, 5 '-конец одной цепи заканчивается рядом с 3'-концом другой, и наоборот. Справа: структура нуклеотида, на которой показана 5'-фосфатная группа и 3'-гидроксильная группа. Обозначения этих групп соответствуют положению на кольце дезоксирибозы. Кольцевые атомы углерода сахара маркированы от 1' (углерод, несущий азотистое основание) до 5' (углерод, несущий фосфатную группу). 3' углерод в середине несет гидроксильную группу. _Изображение изменено, источник: "Химическая структура ДНК," от Madeleine Price Ball (CC0/public domain)._ Правосторонняя спиральВ модели Уотсона и Крика две нити ДНК закручиваются вокруг друг друга, образуя правостороннюю спираль. Все спирали имеют направленность — свойство, показывающее, как их узлы ориентированы в пространстве. [Как определить, что спираль ДНК — правосторонняя?] 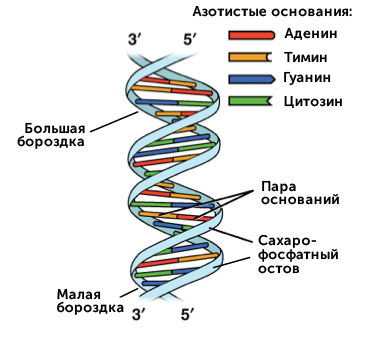 Изображение двойной спирали ДНК, иллюстрирующее её правостороннее закручивание. Большая бороздка представляет собой широкое расстояние между витками одной нити ДНК, в то время как малая бороздка – расстояние между соседними нитями ДНК, и оно намного меньше большой бороздки по размеру. Пары оснований находятся в центре спирали, в то время как сахарнофосфатный остов проходит вдоль внешней стороны. _Изображение изменено, источник: "Структура ДНК и секвенирование: Рисунок 3," от OpenStax College, Biology (CC BY 3,0)._ [Всегда ли спирали ДНК правосторонние?] Геометрическое расположение оснований создают в скрученной спирали более широкий зазор (так называемую большую бороздку) и более узкий зазор (малую бороздку), которые идут вдоль всей молекулы, как показаны на рисунке выше. Эти бороздки являются важными участками связывания с белками, поддерживающими ДНК и регулирующими активность генов. Комплементарность основанийВ модели Уотсона и Крика две нити двойной спирали ДНК удерживаются вместе водородными связями между азотистыми основаниями на противоположных нитях. Каждая пара оснований лежит перпендикулярно нити, образуя «ступеньку» на «лестнице» молекулы ДНК. Пары оснований не состоят из какой-то случайной комбинации оснований. Вместо этого, каждому A находящемуся на одной нити, соответствует T, находящийся на противоположной нити (и наоборот). Точно так же G, находящийся на одной цепи, всегда будет иметь в качестве пары C на противоположной цепи. Эти связи A-T и G-C известны как комплементарные пары оснований. 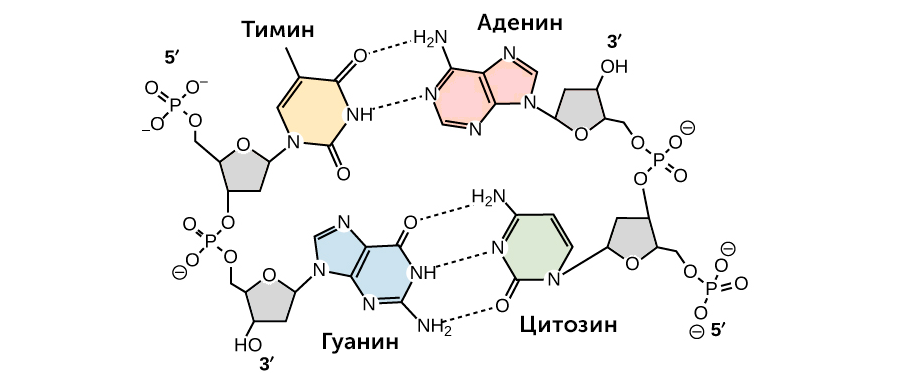 Диаграмма, иллюстрирующая попарное соединение оснований между собой A-T и G-C. А и Т находящиеся напротив друг друга на двух нитях спирали, и их функциональные группы образуют две водородные связи, которые удерживают нити вместе. Точно так же G и C находятся напротив друг друга на двух нитях, и их функциональные группы образуют три водородные связи, которые удерживают цепи вместе. _Изображение изменено, источник: "Структура ДНК и секвенирование: Рисунок 3," от OpenStax College, Biology (CC BY 3,0)._ Парные основания объясняют, почему по правилам Чаргаффа количество A всегда равно количеству T, а количество C — количеству G^{11}11start superscript, 11, end superscript. Когда в одной цепи встречается A, в другой ему обязательно должен соответствовать T, и то же самое с C и G. А поскольку большой пурин (A или G) всегда находится в паре с небольшим пиримидином (T или C), диаметр спирали остаётся постоянным — около 222 нм. Не смотря на то, что первоначальная модель Уотсона и Крика предполагала наличие двух водородных связей между основаниями каждой пары, сегодня мы знаем, что между G и C образуются еще одна дополнительная связь (так, что пары A-T образуют всего две водородные связи, в то время как пары G-C образуют три) ^{12}12start superscript, 12, end superscript. Влияние двойной спиралиСтруктура ДНК открыла дверь для понимания многих аспектов функционирования ДНК, таких как, её копирование и того, как информация, которая содержится в ней, используется клеткой для производства белков. Как мы увидим в следующих статьях и видеороликах, модель Уотсона и Крика дала начало новой эпохе открытий в молекулярной биологии. Эта модель ДНК и основанные на ней последующие открытия составляют основу для большинства современных исследований в области биологии и биомедицины. Дополнительные материалы, помимо материалов Академии ХанаХотите узнать больше о спирали ДНК? Посмотрите эту интерактивную статью от LabXchange. LabXchange - это бесплатная научно-образовательная онлайн-платформа, созданная на факультете искусств и наук Гарварда при поддержке Фонда Амгена. [Ссылки и примечания] Вы студент или |