Книга 2 Издание пятое исправленное и дополненное
 Скачать 5.16 Mb. Скачать 5.16 Mb.
|

14.3. ПИЩЕВАРИТЕЛЬНАЯИ ДЫХАТЕЛЬНАЯ СИСТЕМЫУникальной особенностью организации хордовых является филогенетическая, эмбриональная, а также функциональная связь пищеварительной и дыхательной систем. Действительно, только у хордовых дыхательная система развивается на базе пищеварительной и на первых этапах эволюции функционирует совместно с ней. Так, у ланцетника, сохранившего в значительной степени черты организации предковых форм, специализированных органов дыхания нет, а дыхательную функцию выполняет глотка — передняя часть пищеварительной трубки, пронизанная сквозными отверстиями — жаберными щелями, главной функцией которых является фильтрация воды. Челюстного аппарата у ланцетника нет, и активно питаться он не может. Поэтому источником его питания является взвесь органических частиц в воде, которые задерживаются жаберными перегородками, прилипая к слизи, обильно выделяемой железистыми клетками глотки. Постоянный ток свежей воды через глотку способствует газообмену в кровеносных сосудах, расположенных в жаберных перегородках. На капилляры эти сосуды не распадаются, что свидетельствует о второстепенности дыхательной функции глотки. У более высокоорганизованных хордовых начиная с рыб пищеварительная и дыхательная функции осуществляются специализированными системами, объединенными анатомически общей полостью рта и глотки, а также развитием из общего энтодермального зачатка. 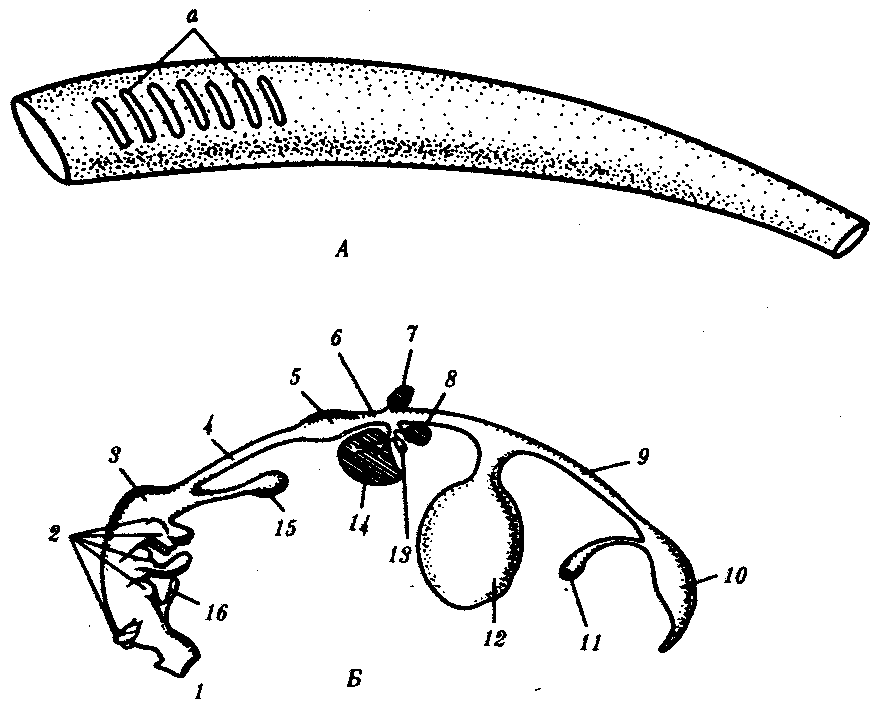 Рис. 14.18. Кишечная трубка позвоночных (А) и ее дифференцировка в эмбриогенезе человека (Б): а—жаберные щели в глотке; 1—ротовая полость, 2—1—5-й глоточные карманы, 3—глотка, 4—пищевод, 5—желудок, 6—двенадцатиперстная кишка, 7—дорсальная поджелудочная железа, 8—вентральная поджелудочная железа, 9— тонкая кишка. 10—клоака, 11—мочевой пузырь, 12—желточный мешок, 13—желчный пузырь, 14—печень, 15—легкие, 16—закладка щитовидной железы Тесная связь обеих систем в филогенезе определяется в первую очередь их топографическими и динамическими координациями, а развитие в онтогенезе — морфогенетическими и эргонтическими корреляциями. Пищеварительная и дыхательная системы хордовых в эмбриогенезе закладываются вначале в виде прямой трубки, подразделяющейся на три участка. Переднюю ее часть, начинающуюся ротовым отверстием и заканчивающуюся переходом в глотку, называют stomodeum. Слизистая оболочка, выстилающая этот участок, эктодермального происхождения и в развитии связана с кожным эпидермисом и его производными (см. § 14.1). Средняя часть кишки начинается глоткой и заканчивается в том месте, где ее энтодермальная слизистая оболочка контактирует с эктодермальной слизистой оболочкой задней кишки, или proctodeum (рис. 14.18). 14.3.1. Ротовая полостьРассмотрим вначале эволюцию ротовой полости и ее производных. У бесчерепных ротовая полость окружена предротовой воронкой со щупальцами и частично выстлана мерцательным эпителием, который вместе с таким же эпителием глотки создает постоянный ток воды в кишечную трубку, несущую пищевые частицы и кислород. Ротовое отверстие позвоночных окружено кожными складками — губами, которые становятся подвижными только у сумчатых и плацентарных млекопитающих в связи со вскармливанием детенышей молоком. Крыша ротовой полости образована у рыб и земноводных основанием мозгового черепа, которое является первичным твердым нёбом. Хоаны земноводных открываются в их ротовую полость сразу позади альвеолярной дуги верхней челюсти. У пресмыкающихся объем ротовой полости увеличивается, и на верхнечелюстных и нёбных костях появляются горизонтальные складки, частично разделяющие ее на верхний, дыхательный, отдел и вторичную ротовую полость. Хоаны при этом несколько смещаются кзади. У млекопитающих наблюдается срастание этих складок по средней линии таким образом, что возникает сплошное вторичное твердое нёбо, полностью отделяющее друг от друга ротовую полость и полость носа. Хоаны при этом открываются в носоглотку. Этим достигается независимость функций органов ротовой полости от процесса дыхания (рис. 14.19, А— В). До 7-й недели эмбрионального развития человека дифференцировка области stomodeum отсутствует. К концу 8-й недели происходит формирование вторичного твердого нёба за счет срастания горизонтальных костных складок. При нарушении адгезии клеток обеих складок возможно незаращение твердого нёба — порок развития, известный под названием волчья пасть (рис. 14.19, Г). Эта аномалия имеет атавистическую природу. Доказаны генетические механизмы ее возникновения. Она сопровождает ряд хромосомных синдромов, а также наследуется изолированно, причем с разной частотой у населения разных популяций. Так, в Японии частота «волчьей пасти» равна 2,1, а в Нигерии — 0,4 случая на 1000 рождений.  Зубы позвоночных связаны по происхождению с плакоидной чешуей хрящевых рыб (см. § 14.1). У них наблюдается непрерывный переход от типичных чешуи к зубам в ротовой полости (рис. 14.20). Зубы первоначально расположены во много рядов и покрывают всю слизистую оболочку ротовой полости, располагаясь у многих рыб даже на языке. У ряда земноводных зубы также расположены не только на альвеолярной дуге, но и на других костях, например на сошнике. У пресмыкающихся обнаруживается только один ряд зубов, причем, так же как у земноводных и рыб, дифференцировка их отсутствует. Такую зубную систему, в которой все зубы одинаковы, называют гомодонтной. У животных перечисленных классов зубы, как и плакоидные чешуи, могут многократно выпадать, сменяясь новыми их поколениями. Многократную смену зубов называют полифиодонтизмом. 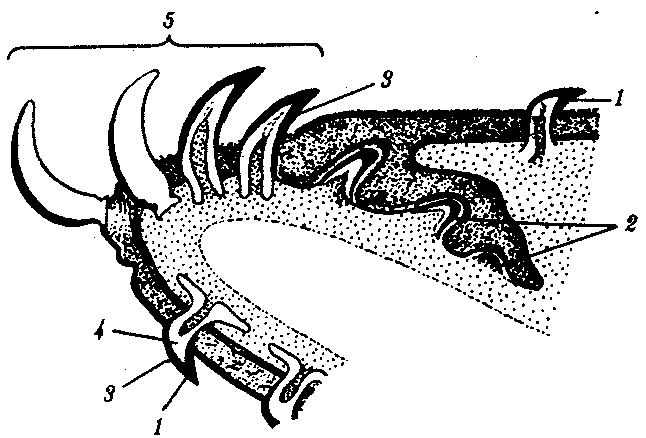 Рис. 14.20. Переход от плакоидной чешуи к зубам по краю ротового отверстия акулы: 1— плакоидная чешуя, 2—закладка новых зубов, 3—эмаль, 4—дентин, 5—зубы Зубы млекопитающих дифференцированы на резцы, клыки и коренные. Они выполняют различные функции. Такую зубную систему называют гетеродонтной. Если резцы, и в особенности клыки, млекопитающих еще весьма сходны с коническими зубами предков, то наибольшим эволюционным преобразованиям подверглись коренные зубы. Впервые они вознили у зверозубых ящеров начала мезозойской эры за счет расширения оснований зубов и появления дополнительных бугорков, а затем и увеличения жевательных поверхностей при их сглаживании. Общее количество зубов у млекопитающих уменьшается и достигает у высших приматов 32. Зубы располагаются только на альвеолярных дугах челюстей, в ячейках. Основание зуба сужается, образуя корень. Зубы человека по сравнению с другими приматами уменьшены в размерах, особенно клыки. Моляры имеют четырехбугорчатое строение. Зубная дуга округлой формы. В связи с дифференцировкой зубов увеличилась продолжительность их функционирования, в результате чего в онтогенезе сменяются только два их поколения: молочные и коренные. Это явление называют дифиодонтизмом. У человека возможны атавистические аномалии зубной системы, связанные с нарушениями как дифференцировки зубов, так и с их количеством. Редкой аномалией является гомодонтная зубная система, в которой все зубы имеют коническую форму. Более частой патологией является трехбугорчатое строение коренных зубов. Нередко встречается прорезывание сверхкомплектных зубов в ряду или за его пределами, иногда даже на твердом нёбе (рис. 14. 21). Это свидетельствует о том, что у человека возможно образование большего количества зубных зачатков, чем 32, как это в норме встречается у низших млекопитающих и представителей более отдаленных классов позвоночных. Свидетельством тенденции к дальнейшему уменьшению количества зубов у человека является то, что нередко последние коренные зубы, так называемые «зубы мудрости», вообще не прорезываются, а если и прорезываются, то это происходит поздно — до 25 лет. Кроме того, эти зубы имеют явно рудиментарный характер, уменьшены в размерах и часто слабо дифференцированы. 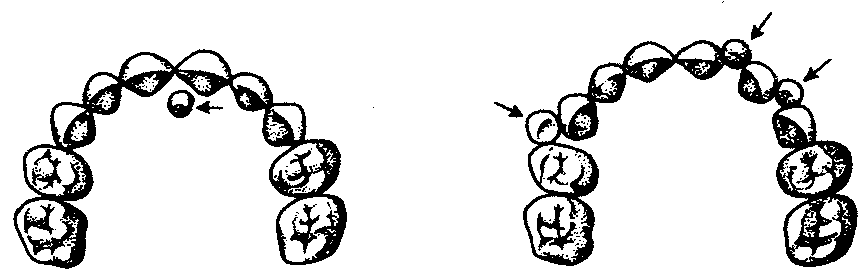 Рис. 14.21. Сверхкомплектные зубы, прорезавшиеся у человека (указаны стрелками) На дне ротовой полости позвоночных располагается непарный выступ — язык, который у рыб представляет собой складку слизистой оболочки, лишенную мышц. Его движения осуществляются вместе с челюстями и подъязычной костью. У наземных позвоночных в языке располагаются мышцы, и они определяют его собственную подвижность. У пресмыкающихся и млекопитающих язык закладывается из трех зачатков: одного непарного и двух парных, лежащих кпереди от первого. Парные зачатки позже срастаются и дают начало телу языка. У большинства рептилий это срастание неполное, и язык раздвоен на конце. Весьма редким пороком развития языка у человека является раздвоенность его конца как результат несращения парных зачатков в эмбриогенезе. Слюнные железы у рыб, заглатывающих пищу вместе с водой, отсутствуют и появляются только при переходе на сушу. Земноводные, обитающие на земле, имеют несколько слюнных желез, выделяющих только слизь. Слюна пресмыкающихся содержит уже и пищеварительные ферменты, а у некоторых змей—и токсины (см. §23.1). У млекопитающих слюнные железы многочисленны: это и мелкие — зубные, щечные, нёбные, язычные, гомологичные железам земноводных и пресмыкающихся, и крупные — подъязычные, подчелюстные и околоушные. Подъязычная и подчелюстная железы — результат дифференцировки подъязычной железы пресмыкающихся, околоушная — новое приобретение млекопитающих. 14.3.2. ГлоткаГлотка — орган, выполняющий у всех хордовых две функции: дыхательную и пищеварительную. У ланцетника она пронизана большим количеством жаберных щелей (более 150 пар). У рыб жаберные щели в количестве 5—7 закладываются как слепые парные выросты глотки — жаберные мешки. Навстречу им выпячиваются кожные покровы — жаберные карманы. В месте их соприкосновения происходит прорыв тканей эктодермы кожи и энтодермы глотки и возникают сквозные жаберные щели. У личинок земноводных в глотке образуется четыре пары жаберных щелей (рис. 14.22), а у пресмыкающихся прорыв их происходит только в эмбриональном развитии; вскоре после этого они зарастают. У млекопитающих в эмбриогенезе начинается закладка глоточных мешков и жаберных карманов, которые в норме никогда не прорываются и не образуют жаберных щелей (рис. 14.23). При нарушении эмбриогенеза во время закладки зачатков жаберных щелей они могут прорываться и даже сохраняться в постэмбриональном периоде у млекопитающих и человека. Эту аномалию называют латеральными свищами шеи. Они открываются на коже боковой поверхности шеи, а другим концом впадают в глотку. Чаще встречается феномен под названием латеральные кисты шеи. Эта аномалия связана с сохранением в постнатальном развитии эмбрионального материала глоточных жаберных мешков или эктодермальных жаберных карманов. Участки этих образований могут заполняться жидкостью, увеличиваться в размерах и склонны к малигнизации. Необходимы своевременная диагностика этих образований и их хирургическое удаление. Первая жаберная щель у всех наземных позвоночных начиная с земноводных в процессе формирования превращается в евстахиеву трубу, барабанную полость и наружный слуховой проход. Располагаясь топографически в зоне редукции первичных челюстей и подъязычной висцеральной дуги, преобразующихся в слуховые косточки, она меняет свои функции, включаясь в систему слухового анализатора (см. разд. 14.2.1).  Рис. 14.22. Закладка жаберных щелей у позвоночных. А—фронтальный срез личинки тритона; Б—срез через глотку зародыша мыши (головной конец зародыша сильно изогнут, поэтому на фотографии видны два среза одной нервной трубки): 1—5—жаберные карманы и жаберные щели, 6—зачаток легкого, 7—полость глотки, 8—нервная трубка 14.3.3. Средняя и задняя кишкаКишечная трубка претерпевает в ходе филогенеза следующие прогрессивные преобразования: увеличение общей длины, дифференцировку на отделы и образование крупных многоклеточных желез. Если длина этого отдела пищеварительной трубки ланцетника составляет примерно 1/3 длины тела, то у млекопитающих возможно превышение ее длины по отношению к телу в 10 раз и более. Этим достигается удлинение времени контакта пищевых частиц с ферментами пищеварения и увеличение всасывающей поверхности. На органном уровне это сопровождается появлением складчатости слизистой оболочки кишки, возникновением ворсинок и крипт. Кишечник ланцетника абсолютно не дифференцирован. В средней его части расположен печеночный вырост — простое слепое выпячивание брюшной стенки кишки, выстланное железистыми клетками. Функционально этот вырост представляет собой зачаточную печень. Он является единственной многоклеточной железой пищеварительной системы ланцетника. 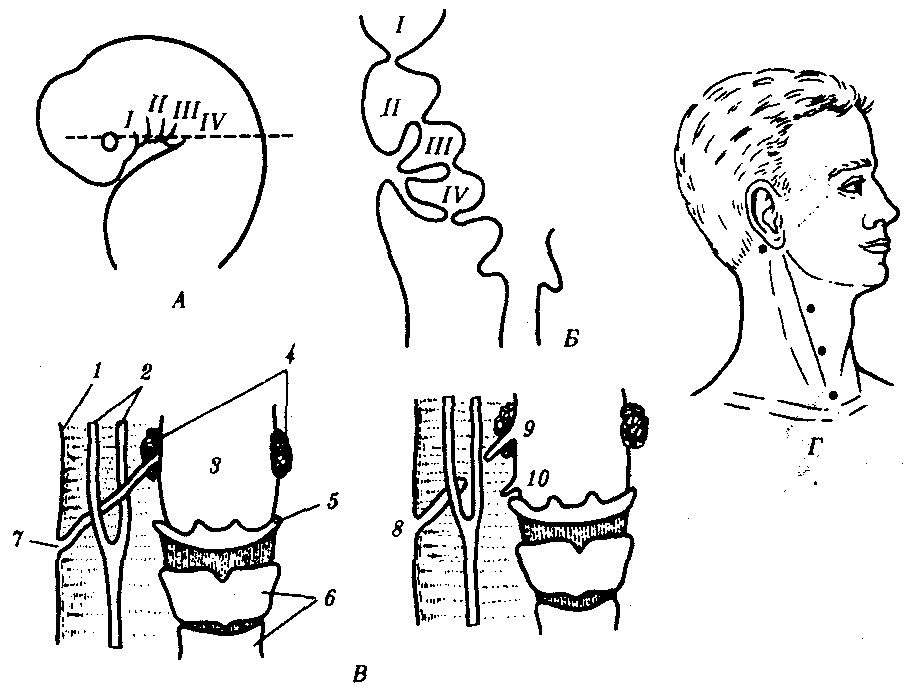 Рис. 14.23. Развитие области глотки и ее аномалии у человека. А—зародыш в возрасте 5 недель; Б—тот же зародыш (срез через область глотки по пунктирной линии, изображенной на рис. А); В—схема латеральных свищей шеи; Г—проекции наиболее часто встречающихся латеральных шейных свищей: I—IV—жаберные дуги; 1—кожа, 2—сонная артерия, 3—глотка, 4—нёбные миндалины, 5—подъязычная кость, 6—гортань, 7—сквозной шейный свищ, 8—10—несквозные свищи У рыб за глоткой следует короткий пищевод, затем желудок, слабо от него отграниченный. В кишечнике выделяются тонкий и толстый отделы, последний открывается анусом во внешнюю среду. В отличие от ланцетника у рыб хорошо развита печень, снабженная желчным пузырем. Поджелудочная железа у различных рыб построена по-разному. В некоторых случаях она представлена отдельными мелкими дольками в стенке кишечника, в мезентерии, рассеяна в ткани печени. Нередко уже у рыб она представляет собой компактный орган, включающий в себя как экзокринную часть, которая ответственна за синтез пищеварительных ферментов, так и эндокринную, которая выделяет гормоны, регулирующие углеводный обмен. Таким образом, у рыб наблюдаются все основные стадии эволюции сложной многоклеточной железы, структура и функции которой впоследствии принципиально практически не изменяются. Единственное отличие пищеварительной трубки земноводных от трубки рыб — ее удлинение и впадение толстой кишки в клоаку. Существенной особенностью пищеварительной трубки пресмыкающихся является возникновение в ней слепой кишки. У большинства из них она зачаточна, но у некоторых развита весьма существенно. Слепая кишка — важное эволюционное приобретение пресмыкающихся, позволяющее им расширить рацион питания и использовать растительную пищу, переваривание которой сложно и требует участия симбиотических простейших и бактерий. Слепая кишка особенно богата микрофлорой, под действием которой осуществляются процессы брожения, позволяющие наиболее полно использовать пластические вещества растительного происхождения. У млекопитающих за счет дальнейшей дифференцировки пищеварительного тракта, и в частности больших размеров слепой кишки, резко увеличивается разнообразие используемых продуктов питания и возникает пищевая специализация на травоядность и плотоядность; всеядность встречается редко, в частности в отряде приматов. Задняя кишка плацентарных млекопитающих дифференцирована, клоака отсутствует и прямая кишка заканчивается анусом. В онтогенезе человека рекапитулируют основные стадии филогенеза кишечной трубки и ее производных. Из пороков развития, связанных с задержкой эмбриогенеза, известны гипоплазии всей пищеварительной системы, укорочение кишки и недоразвитие любых ее отделов, а также печени и поджелудочной железы. В зависимости от степени недоразвития тяжесть аномалии может быть как незначительной, так и несовместимой с жизнью. Клинический интерес представляет гетеротопия тканей поджелудочной железы в стенке тонкого кишечника или желудка. Иногда гетеротопированные фрагменты могут симулировать опухолевую трансформацию слизистой оболочки. По данным патологоанатомов, частота этой аномалии — от 0,6 до 5%. Механизм гетеротопии — нарушение клеточной миграции зачатков железы из стенки кишечной трубки. С недоразвитием и задержкой дифференцировки связана и такая аномалия, как персистирование клоаки, при которой мочеполовые пути и прямая кишка объединены. При нормальном развитии после 8-й недели эмбриогенеза клоака должна полностью дифференцироваться на прямую кишку, мочевыделительные и половые протоки. 14.3.4. Органы дыханияЖабры — наиболее ранние специализированные органы дыхания, появляющиеся среди хордовых впервые у рыб. Они представляют собой тонкие складки слизистой оболочки глотки, лежащие на жаберных дугах, снабжаемые венозной кровью через жаберные артерии и распадающиеся здесь на капилляры. Позади последней жаберной дуги у кистеперых рыб за счет выпячивания вентральной стороны глотки формируется парное образование — плавательный пузырь, который выполняет в первую очередь гидростатические функции, уравновешивая тело рыбы в толще воды. Связь между ним и глоткой, имеющаяся в эмбриогенезе, не прерывается и позже, поэтому воздух при попадании в глотку может беспрепятственно проходить в плавательный пузырь. Эта морфофизиологи-ческая особенность при изменении условий существования кистеперых рыб явилась предпосылкой преобразования плавательного пузыря в легкие (рис. 14.24). 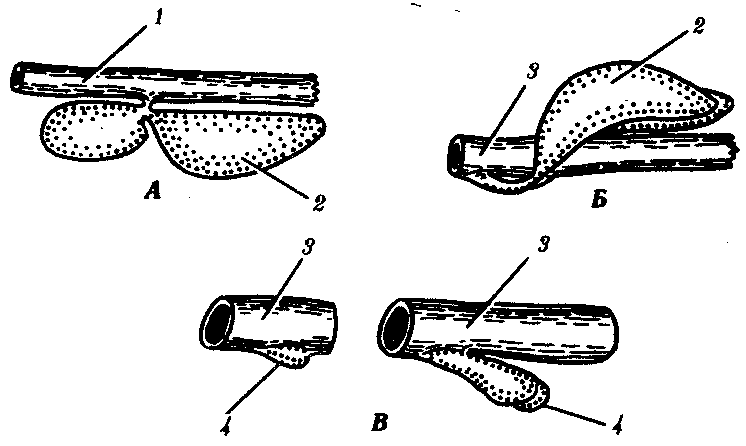 Рис. 14.24. Плавательный пузырь рыб (А—костной; Б—кистеперой) и развитие легких у человека (В—ранние стадии): 1—средняя кишка, 2—плавательный пузырь, 3—глотка, 4—развивающиеся легкие У земноводных, связанных по происхождению с кистеперыми рыбами, в личиночном состоянии функционируют жабры, а во взрослом — органами дыхания становятся легкие. В связи с отсутствием грудной клетки и диафрагмы воздух в них попадает из ротовой полости за счет глотательных движений, осуществляющихся подбородочно-подъязычной мышцей. Материал жаберных дуг, следующих за подъязычной дугой, частично редуцируясь, входит в состав хрящей гортани, которая, появляясь впервые у земноводных, является первым органом, относящимся к нижним дыхательным путям. Легкие начинаются непосредственно от гортани. Они крупноячеисты и имеют относительно малую дыхательную поверхность, в связи с чем газообмен в большей степени осуществляется через кожные покровы (см. § 14.1). Пресмыкающиеся, вышедшие на сушу окончательно, имеют как верхние дыхательные пути (не полностью отграниченную от ротовой полости полость носа), так и нижние — гортань, трахею и бронхи. Их легкие мелкоячеисты, содержат многочисленные внутренние перегородки и обладают большой дыхательной поверхностью. Впервые появляется диафрагма, которая в дыхании принимает скорее пассивное участие, так как она либо лишена мышечных элементов, либо разделяет грудную и брюшную полости не полностью. Механизм дыхания основан на сокращении межреберных мышц, приводящих в движение грудную клетку. У млекопитающих дыхательные пути, как и у предков, выстланы мерцательным эпителием. Они полностью отделены от пищеварительной системы и только перекрещиваются с ней в глотке. Бронхи многократно разветвляются, вплоть до бронхиол, ведущих в альвеолы — легочные пузырьки, имеющие в сумме огромную площадь поверхности (у человека до 90 м ). Основной мышцей, изменяющей объем грудной полости, становится диафрагма. В эмбриогенезе человека отражается первоначальное единство пищеварительной и дыхательной систем. На этом основано формирование большой группы врожденных пороков развития пищевода и трахеи типа эзофаготрахеальных свищей (рис. 14.25). Нарушением альвеолярной дифференцировки легочной ткани могут быть объяснены дизонтогенетические бронхолегочные кисты — округлые полости в легких, отграниченные от окружающих тканей примитивно построенной стенкой недифференцированного бронха, а также кистозная гипоплазия (недоразвитие) легких (рис. 14.26). При этой патологии недоразвитыми оказываются целые доли легкого, которые представляют собой многочисленные полости, связанные с крупными бронхами и имеющие малую поверхность. Газообмен в таких участках легких резко ослаблен. Широко известны также гипоплазии диафрагмы от небольших дефектов в ее куполе до полной аплазии. Последняя аномалия несовместима с жизнью и встречается обычно вместе с другими множественными пороками развития. 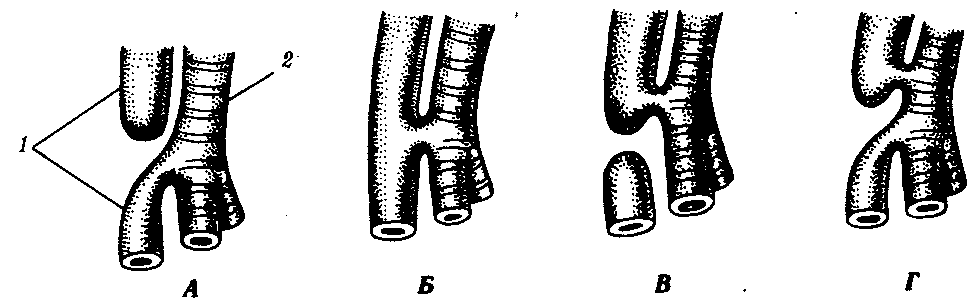 Рис. 14.25. Различные формы эзофаготрахеальных свищей (А—Г): 1— пищевод, 2— трахея |