Книга 2 Издание пятое исправленное и дополненное
 Скачать 5.16 Mb. Скачать 5.16 Mb.
|

ГЛАВА 15АНТРОПОГЕНЕЗИ ДАЛЬНЕЙШАЯ ЭВОЛЮЦИЯ ЧЕЛОВЕКА15.1. МЕСТО ЧЕЛОВЕКАВ СИСТЕМЕ ЖИВОТНОГО МИРАНеограниченный прогресс в эволюции живой материи проявился в возникновении человека как биосоциального существа. Появление человека с его целенаправленной трудовой деятельностью качественно изменило облик планеты Земля и определило новое направление эволюции природы в целом. Будучи биологическим видом, человек — единственный на Земле организм, обладающий социальной сущностью. Социальная сущность человека определяет его современное состояние и прогнозирует будущее. Биологическое в нем при этом становится его наследством, поэтому эволюция человека на современном этапе идет не столько по биологическим законам, сколько по законам развития человеческого общества. Биологическая эволюция представляет собой биологическую форму движения материи, а эволюция общества — социальную. Естественно, что социальная форма движения материи возникла лишь с появлением человека. Поэтому изучение вопросов антропогенеза — это не только биологическая, но и философская проблема. Человек как биологический вид относится к отряду приматов (рис. 15.1). 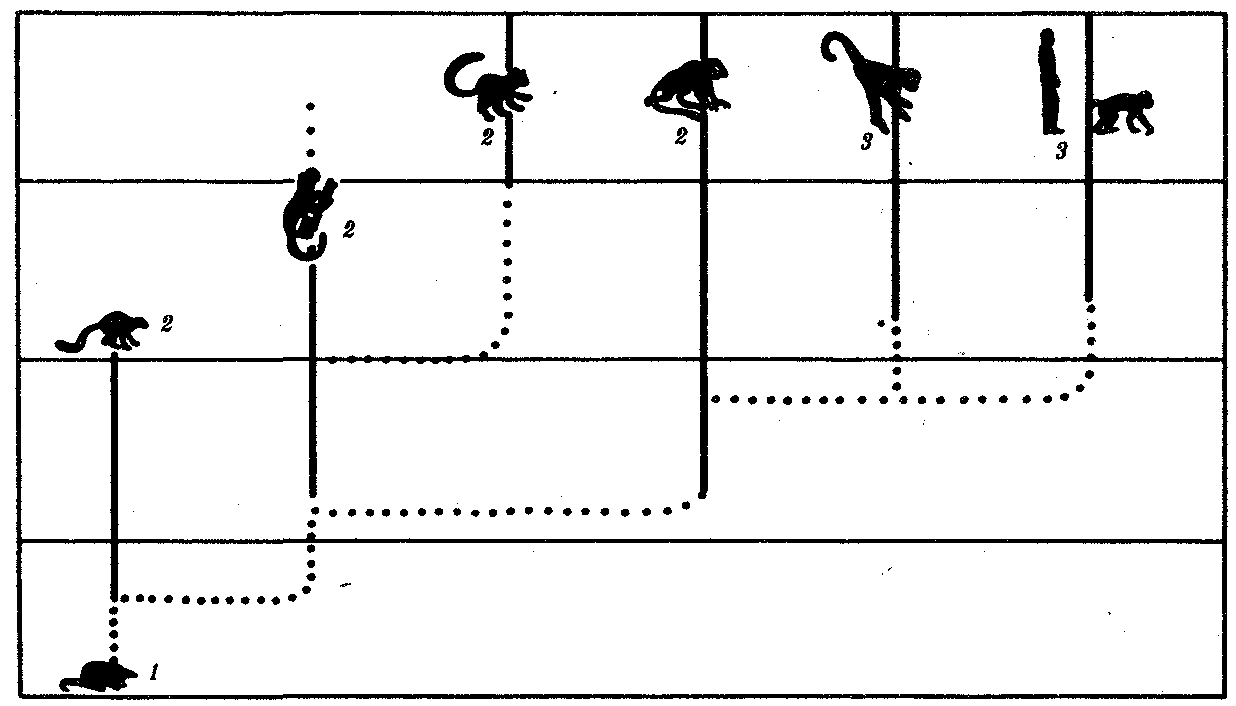 Рис. 15.1. Место человека в отряде приматов: 1—предок приматов, 2—вымершие и современные полуобезьяны, 3— настоящие обезьяны Нового, Старого света и человек С биологических позиций важно ответить на вопрос: почему человек возник именно в отряде приматов, а не в другом эволюционно прогрессивном отряде млекопитающих. Ключевой адаптацией представителей отряда приматов являются древесный образ жизни и передвижение по ветвям с помощью хватательных конечностей, имеющих противопоставленный большой палец и уплощенные ногти вместо когтей. Пальцевые подушечки и внутренние поверхности кистей и стоп имеют кожные узоры в виде гребешков и бороздок, увеличивающих контакт пальцев с ветками и усиливающих их тактильную чувствительность. Локтевая кость свободно вращается вокруг лучевой. Эти особенности при освобождении рук от участия в движении являлись предпосылкой к использованию их в трудовой деятельности. Другими прогрессивными чертами приматов, связанными с энергичными перемещениями в трехмерном пространстве, являются бинокулярное зрение, сильно развитые мозжечок и кора больших полушарий головного мозга. В отличие от большинства отрядов млекопитающих приматов характеризует также выраженная социальность: жизнь небольшими стадами со сложной системой иерархии и общественным воспитанием потомства. Плодовитость приматов чрезвычайно низка — обычно рождается один детеныш, беспомощный и нуждающийся в уходе на протяжении многих лет. Характерной особенностью поведения приматов является выраженный инстинкт подражания, обеспечивающий быстрое научение и возможность передачи социального опыта в ряду поколений. Перечисленные черты отряда приматов явились необходимыми преадаптациями к возникновению человека при появлении комплекса благоприятных условий. 15.2. МЕТОДЫ ИЗУЧЕНИЯЭВОЛЮЦИИ ЧЕЛОВЕКАОбычно выделяют следующие этапы эволюции человека: 1. Древнейшие стадии гоминизации — происхождение рода Homo. 2. Эволюция рода Homo до возникновения современного человека. 3. Эволюция современного человека. Первый этап антропогенеза есть чисто биологическая эволюция. На втором этапе к элементарным факторам биологической эволюции подключается действие социального фактора, который на третьем этапе является доминирующим. Методологические подходы к изучению разных этапов антропогенеза различны. Изучение 1-го этапа производят методами палеонтологии и сравнительной анатомии. В связи с появлением элементов материальной культуры 2-й этап изучают также методами археологии. На 3-м этапе эволюционные события происходят в основном на молекулярно-генетическом уровне и проявляются на популяционном уровне. Основными способами изучения эволюции современного человека поэтому являются биохимический, цитогенетический и популяционно-статистический методы. Изучение первых двух этапов эволюции человека сталкивается с необходимостью датировки палеонтологического материала и элементов материальной культуры. Для определения абсолютного возраста ископаемых остатков человека и его предков широко используют физические методы, в частности радиометрические. С помощью масс-спектрометров определяют изотопный состав изучаемого объекта и по соотношению элементов с учетом периода полураспада входящих в его состав радиоактивных изотопов выявляют возраст образца. Ископаемый костный материал содержит в своем составе минеральные компоненты и белок коллаген, разрушающийся чрезвычайно медленно. На этом основан коллагеновый метод абсолютной датировки ископаемых остатков: чем меньше коллагена содержится в образце, тем более велика его древность. В последнее время в антропологии все более активно применяют методы иммунологии, молекулярной биологии и цитогенетики. В связи с огромной важностью этих методов остановимся на них подробнее. Для определения прямого родства организмов друг с другом используют иммунологический метод, основанный на изучении иммунологических реакций антиген — антитело. Его можно применять для изучения степени родства не только современного человека с человекообразными обезьянами, но и ныне живущих видов с ископаемыми. Для этого следовые количества белка, извлекаемые из костей ископаемых форм, используют для получения антител, которые и применяют в иммунных реакциях с белками современных видов. Из современных человекообразных обезьян к человеку иммунологически наиболее близок шимпанзе, наиболее далеко от человека отстоит орангутан. Иммунологическим методом было обнаружено, что белки рамапитека, человекообразной обезьяны Южной Азии (абс. возраст 13 млн. лет), более сходны с белками орангутана, чем человека и шимпанзе. Эти данные вместе с результатами морфологических и палеонтологических сопоставлений заставили отказаться от представления о том, что рамапитек является прямым предком человека, и связать его с эволюционной линией орангутана. Из этого следует, что разделение человеческой линии эволюции с африканскими человекообразными обезьянами произошло значительно позже, чем 13 млн. лет назад. В последние годы антропогенез эффективно изучают также биомолекулярными методами. В основе этих методов изучения эволюции лежит допущение, что мера сходства двух таксонов соответствует мере их родства. Поэтому организмы, имевшие общего предка в недалеком прошлом, будут более'сходными друг с другом, чем имевшие его очень давно. Сущность биомолекулярных методов состоит в использовании данных, полученных при сопоставлении белков и нуклеиновых кислот организмов разных видов для определения их родства и древности соответствующих филогенетических ветвей. При этом считают, что степень различий в аминокислотном составе белков и в нуклеотидных последовательностях ДНК позволяет судить о времени расхождения сравниваемых видов от предковой формы. Естественно, что оценивать степень родства и давность расхождения филогенетических ветвей можно лишь по накоплению нейтральных мутаций в геноме, так как вредные мутации быстро элиминируются из генофондов. В действительности нельзя считать большинство мутаций селективно нейтральными. Расчеты показывают, что при нейтральности мутаций скорость их накопления постоянна только при измерении времени в количестве поколений, а не в годах. Так как продолжительность жизни поколений у различных видов различна, то и скорости накопления различий нуклеотидных последовательностей будут более велики у короткоживущих видов по сравнению с долгоживущими. Кроме того, фактическая скорость эволюции может значительно варьировать в различные временные интервалы в различных группах и по разным признакам. В определении применимости методов молекулярной биологии имеет значение возможность возникновения конвергентного сходства молекул, причем вероятность его повышается с увеличением изучаемых временных интервалов. Эволюция генов и белков часто может опережать реальное расхождение популяций, в первую очередь за счет адаптивного генетического полиморфизма. Однако молекулярно-биологические методы применимы для оценки родства и времени дивергенции в качестве приблизительных «молекулярных часов» при сравнении средних скоростей замен нуклеотидов в ДНК в целом и аминокислот во многих белках за длительные интервалы времени. Биомолекулярный подход — лишь один из путей определения эволюционных расстояний, который работает только наряду с классическими методами палеонтологии и антропологии, причем в результатах при этом возможны серьезные расхождения. Так, при изучении скелета человека, обнаруженного на территории Эквадора, по данным радиоуглеродного и аминокислотного анализа была установлена его древность в 28 тыс. лет. При использовании же коллагенового анализа возраст того же скелета оказался датированным не более чем в 2,5 тыс. лет. В антропологии применяют несколько методов оценки степени дифференцированности таксонов, дополняющих друг друга: гибридизация ДНК, определение аминокислотных последовательностей белков, изучение генов гистосовместимости и т.д. Наиболее достоверные данные получены путем гибридизации ДНК, так как скорость эволюции ДНК в целом более постоянна, чем скорость изменения белков. Гибридизация ДНК показала, что момент дивергенции эволюционных ветвей человека и шимпанзе наступил 6,5—6,7 млн. лет назад (рис. 15.2). Однако между эволюцией структуры генома в виде накопления генных мутаций и морфофизиологической эволюцией часто нет прямой зависимости (рис. 15.3). Это может быть связано с тем, что в формировании практически всех сложных фенотипических признаков принимают участие различные генные системы. Таким образом, скорость эволюции белков у двух разных родственных видов может быть одинакова, а скорость эволюции в целом, оцененная по комплексу фенотипических признаков, при этом оказывается различной. Сравнение аминокислотных последовательностей белков шимпанзе и человека привело к выводу, что около 99% их белков абсолютно идентичны. Из этого следует, что и структурные гены человека и шимпанзе сходны в наибольшей степени. С чем же связаны столь значительные морфофизиологические отличия обоих видов? Можно предполагать, что это зависит от различного распределения белков в клетках организма в процессе развития, что, в свою очередь, определяется различиями программы считывания сходной наследственной информации во времени и пространстве. Иными словами, это может быть обусловлено эволюционными преобразованиями в большей степени не структурных, а регуляторных генов. 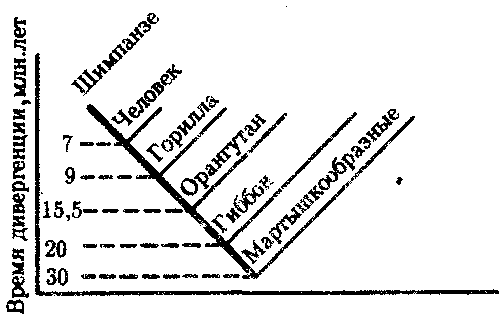 Рис. 15.2. Среднее время дивергенции высших приматов по часам ДНК 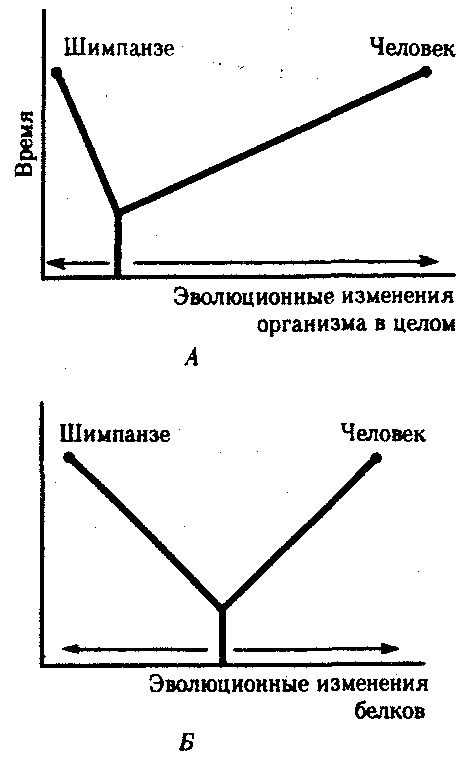 Рис. 15.3. Соотношение скорости морфофизиологической (А) и молекулярной (Б) эволюции Поскольку изучение хромосомного материала возможно только у ныне живущих организмов, применение цитогенетического метода ограничивается современным человеком и человекообразными обезьянами. Дифференциальная окраска хромосом позволяет не только сопоставлять хромосомы разных видов приматов и человека и изучать хромосомный полиморфизм современного человека, но и решать некоторые вопросы эволюции. Выяснено, что кариотип человекообразных обезьян отличается по количеству хромосом от кариотипа человека на одну пару (23 пары хромосом человека и 24 пары шимпанзе). У человека и шимпанзе идентичны 13 пар хромосом. Хромосома 2-й пары человека точно соответствует двум соединенным хромосомам шимпанзе, а остальные хромосомы отличаются друг от друга незначительно. Так, хромосома 5-й пары шимпанзе соответствует такой же хромосоме человека, но небольшой ее перицентрический участок инвертирован на 180° по сравнению с человеческой хромосомой. Инверсии такого рода обнаружены в кариотипах человека и шимпанзе еще в восьми хромосомах. Эти данные вместе с указаниями на сходство белков человека и шимпанзе свидетельствуют об их значительной эволюционной близости. Сопоставление кариотипов людей, происходящих из разных популяций, приводит к выводу о полиморфизме хромосом, в первую очередь по размерам гетерохроматиновых участков. Наследуемость индивидуальных вариаций хромосом и их неравномерное распределение в разных популяциях (в частности, расовые различия по размерам длинного плеча Y-хромосомы) делают возможным популяционно-цитогенетический подход в изучении эволюции современного человека. 15.3. ХАРАКТЕРИСТИКА ОСНОВНЫХ ЭТАПОВ АНТРОПОГЕНЕЗАДолгое время многие палеонтологи и антропологи считали наиболее древними представителями семейства гоминид южно-африканских двуногих приматов Australopithecus africanus. Возраст большей части южно-африканских находок австралопитеков определяется в интервале 3—1 млн. лет. Строение скелетов этих организмов свидетельствует об их прямохождении. Масса мозга — 450—550 г при общей массе тела от 25 до 65 кг. Зубная система близка к человеческой: клыки малых 146 размеров, зубы расположены в виде широкой дуги, как у человека. Это свидетельствует о всеядности. В местах обнаружения остатков австралопитеков имеется множество костей, расколотых тяжелыми предметами. Большое число черепов животных расколоты с левой стороны, и это свидетельствует о том, что австралопитеки были в основном правшами. Некоторые австралопитеки, видимо, начинали осваивать огонь. В 1960—1970-х гг. в результате раскопок в Эфиопии и позже в Танзании и Кении были обнаружены большие группы гоминид, древность которых определяется в 4—2,8 млн. лет. По этим материалам в 1978 г. был описан новый вид австралопитеков A. afarensis, более примитивный, чем A. africanus, но тоже двуногий и с чертами «промежуточности» по отношению к современным человекообразным обезьянам и человеку (рис. 15.4). Объем мозговой полости черепа оказался в пределах 380—450 см3, т.е. практически не отличался от объема мозга современного шимпанзе.  Рис. 15.4. Австралопитек афарский. Реконструкция Действительно, австалопитеки имеют большее сходство с человеком не по объему и строению мозга, а по строению зубной системы и способу передвижения. Изучение австралопитековых показало, что именно двуногость, а не большой объем мозга, явилась ключевой адаптацией ранних гоминид. В 1964 г. по находкам, сделанным в Танзании, был выделен вид Homo habilis, или Человек умелый, имеющий абсолютный возраст 2—1,7 млн. лет. Отличительными чертами его являются двуногость, в целом прогрессивное строение кисти, зубной системы, объем мозговой коробки от 540 до 700 см3, что примерно в полтора раза превышает объем мозга австралопитеков. На внутренней поверхности черепа обнаруживаются признаки прогрессивных нейроморфологических изменений, определяющиеся по отпечаткам головного мозга: выраженная асимметрия полушарий и развитие двух речевых центров как условие для возникновения членораздельной речи. Большой палец стопы не отведен в сторону. Это свидетельствует о том, что морфологические перестройки, связанные с прямохождением, у него полностью завершились. Вместе с остатками Н. habilis найдены орудия труда со следами целенаправленной обработки, свидетельствующие о ранних формах трудовой деятельности. Перечисленные признаки, ведущим из которых является прогрессивное развитие мозга, характеризуют организм уже с иной морфофункциональной организацией по сравнению с австралопитеками. Эти признаки считаются специфичными для рода Homo. Сопоставление морфологии африканского и афарского австралопитеков с Н. habilis и современным человеком позволяет предположить следующую схему филогенетических взаимоотношений ранних гоминид (рис. 15.5). Африканский австралопитек, считавшийся раньше общим предком как рода Homo, так и других австралопитековых, является в этой схеме представителем боковой ветки эволюции, приведшей к узкой специализации и образованию мощных форм типа A. robustus, которые вымерли около 1 млн. лет назад. Общим предком всех австралопитековых и рода Homo является в соответствии с этой схемой A. afarensis. 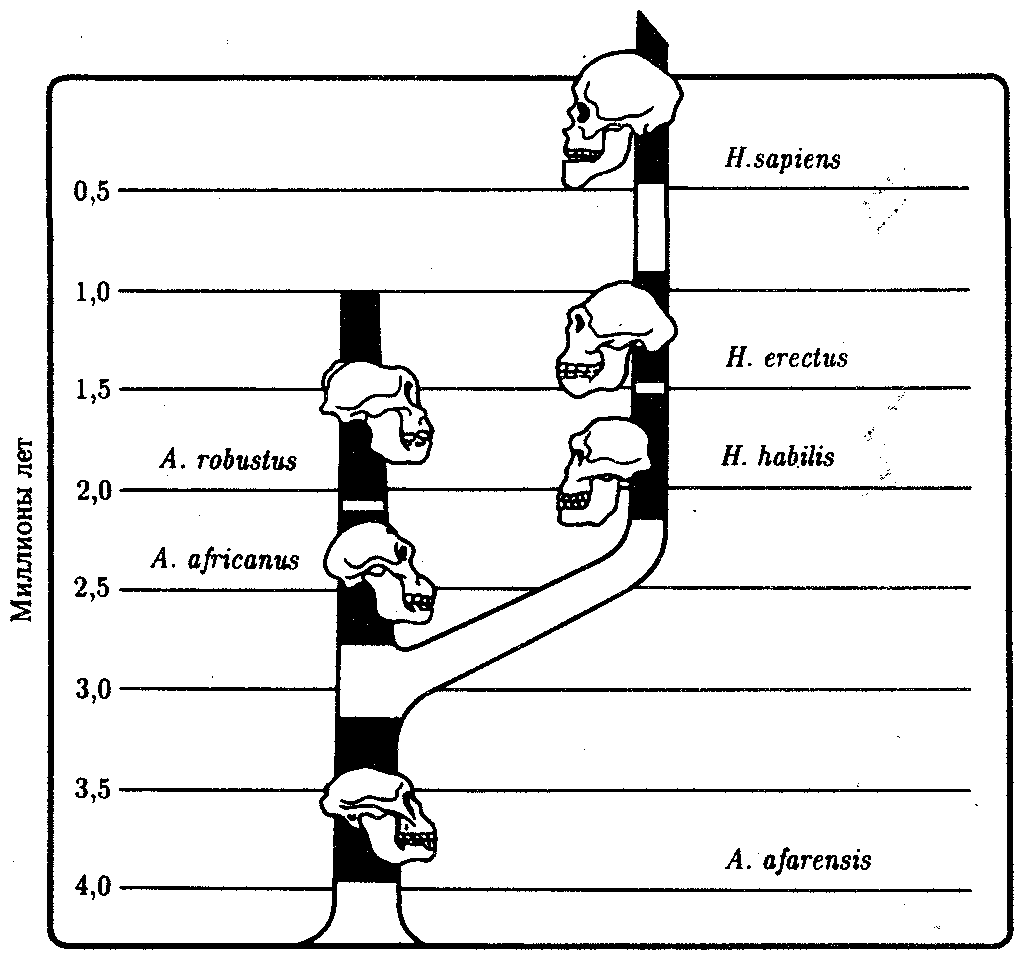 Рис. 15.5. Филогенетические взаимоотношения ранних гоминид Таким образом, на протяжении 1—1,5 млн. лет представители двух близких родов и, возможно, нескольких видов семейства гоминид сосуществовали, причем не только во времени, но и на перекрывающихся территориях. В основе дивергенции различных линий ранних гоминид и австралопитековых могли лежать разного рода механизмы изоляции, в первую очередь генетические: мутации в виде хромосомных перестроек. Это означает, что эволюция австралопитековых шла постепенно, приводя благодаря дивергенции к морфологическому и экологическому разнообразию. Ведущими факторами эволюции на прегоминидной стадии антропогенеза являлись, несомненно, факторы биологической эволюции, главным из которых является естественный отбор. Об этом свидетельствует, в частности, большое видовое разнообразие австралопитековых, обитавших в различных условиях практически на всей территории Южной, Центральной и Северо-Восточной Африки. В это же время в происхождении рода Homo имело место скачкообразное изменение наследственного материала. В разных органах и системах прегоминид обнаруживалась асинхронность филогенеза. Есть предположение о том, что эволюция коры больших полушарий мозга состоит из двух компонентов, разобщенных по времени: соматического, обеспечивающего сенсорно-моторные функции, и несоматического, связанного с высшими психическими функциями. Если локомоторный комплекс подвергался длительным постоянным изменениям, то головной мозг эволюционировал скачкообразно. Элементы скачкообразности в эволюции некоторых структур ранних гоминид могли быть обусловлены «транспозиционными взрывами» (перестройками генома за счет подвижных генетических элементов, переносящих комплексы генов на новое место). Это могло сопровождаться постепенным развитием других морфофизиологических свойств в результате накопления малых мутаций под контролем естественного отбора. Но именно в период становления Н. habilis возникла, вероятно, часть хромосомных перестроек в геноме человека, о которых говорилось выше. Следующей ступенью гоминизации после появления Н. habilis считается возникновение архантропов, представителем которых является сборный вид Н. erectus, или Человек прямоходящий. Материальная культура и ярко выраженная социальность позволили ему быстро и эффективно расселиться по всей территории Африки и Евразии и освоить обширный ареал, разнообразный в климатическом отношении. Действительно, орудия труда Н. erectus более прогрессивны, чем у Н. habilis, а масса мозга (от 800 до 1000 г) явно превышает минимальную массу (750 г), при которой возможно существование речи. Наличие при этом речевых центров, возникших впервые у Н. habilis, предполагает и развитие второй сигнальной системы. Выделяют три группы Н. erectus: обитавшие в Европе, Азии и Африке. Долгое время древнейшими архантропами считались азиатские представители из Индонезии и Восточного Китая — питекантроп и синантроп. Однако находки последних лет на территории Израиля (1982) и Кении (1984), датирующиеся соответственно 2,0 и 1,6 млн. лет, сопровождающиеся элементами материальной культуры и признаками использования огня, показали, что арогенная эволюция гоминид происходила на африканском континенте и на Ближнем Востоке. Это позволило связать происхождение Н. erectus с восточноафриканскими формами Н. habilis. Наличие большого количества находок архантропов древностью 1,5—0,1 млн. лет в отдаленных от Африки регионах — в Юго-Восточной и Восточной Азии, в Центральной Европе и даже на Британских о-вах — свидетельствуют об активных адаптациях их к разнообразным условиям существования (рис. 15.6). В связи с тем что небольшое различие ископаемых остатков Н. erectus не соответствует значительному разнообразию природно-климатических условий указанных территорий, можно заключить, что в этих адаптациях значительную роль играли наряду с факторами биологической эволюции также и социальные факторы: совместное изготовление укрытий, орудий труда и использование огня. 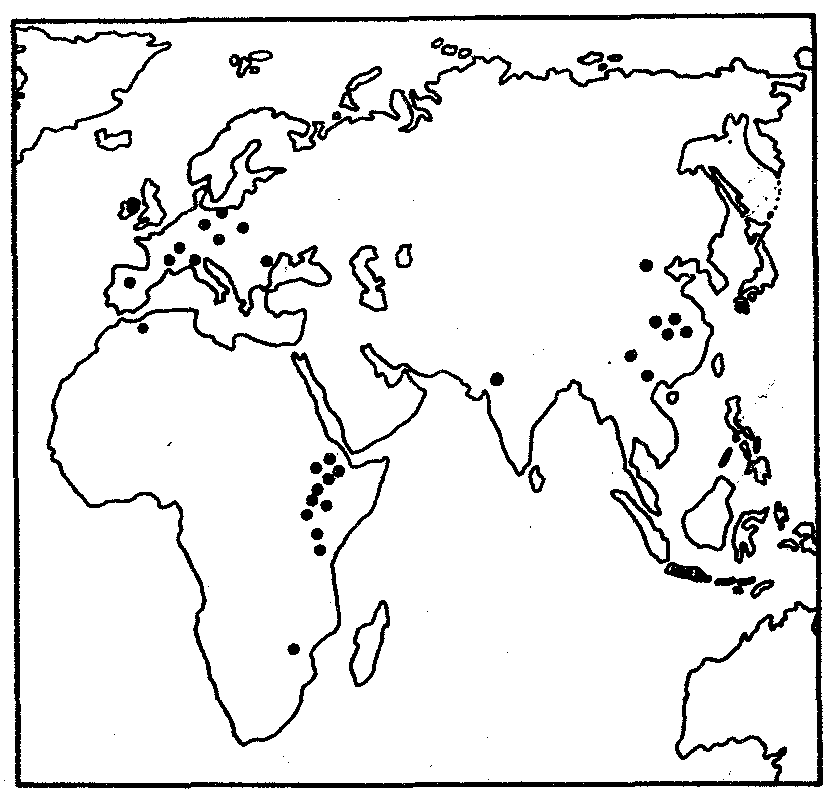 Рис. 15.6. Места обнаружения ископаемых остатков Человека прямоходящего Роль Н. erectus как этапа антропогенеза никогда не подвергалась сомнению. Что же касается палеоантропа, или неандертальского человека, то его роль в происхождении человека в настоящее время оспаривается. Это связано в первую очередь с обнаружением большого количества ископаемых остатков человека с промежуточными чертами между Н. erectus и человеком современного физического типа. Кроме того, палеонтологические находки последних лет дают повод судить о недооценке интеллектуальных возможностей неандертальцев. На всех стоянках обнаружены следы костров и обгоревшие кости животных, что свидетельствует об использовании огня для приготовления пищи. Орудия труда их гораздо совершеннее, чем у предковых форм. Масса мозга неандертальцев около 1500 г, причем сильное развитие получили отделы, связанные с логическим мышлением. Костные остатки неандертальца из Сен-Сезер (Франция) были найдены вместе с орудиями труда, свойственными верхнепалеолитическому человеку, что свидетельствует об отсутствии резкой интеллектуальной грани между неандертальцем и современным человеком. Имеются данные о ритуальных захоронениях неандертальцев на территории Ближнего Востока. Эти и ряд других находок позволили в конце 60-х годов выделить палеоантропов в отдельный подвид Homo sapiens neanderthalensis в отличие от неоантропа H.s. sapiens, который, таким образом, тоже получил ранг подвида. Наиболее древние ископаемые остатки его возрастом 100 тыс. лет обнаружены также на территории Северо-Восточной Африки. Многочисленные находки палеоантропов и неоантропов на территории Европы, датирующиеся 37—25 тыс. лет, свидетельствуют о существовании обоих подвидов в течение нескольких тысячелетий. В тот же период неоантропы обитали уже не только в Европе и Африке, но и в отдаленных районах Азии (о. Тайвань, о. Окинава) и даже в Америке. Эти данные указывают на необычайно быстрый процесс расселения современного человека, что может быть доказательством «взрывного», скачкообразного характера антропогенеза в этот период как в биологическом, так и в социальном смысле. H.s. neanderthalensis в виде ископаемых остатков не обнаруживается позже рубежа в 25 тыс. лет. Быстрое исчезновение палеоантропов может быть объяснено вытеснением их людьми с более совершенной техникой изготовления орудий труда и метисацией с ними. С возникновением человека современного физического типа роль биологических факторов в его эволюции свелась к минимуму, уступив место социальной эволюции. Об этом отчетливо свидетельствует отсутствие существенных различий между ископаемым человеком, жившим 30—25 тыс. лет назад, и нашим современником. 15.4. ВНУТРИВИДОВАЯ ДИФФЕРЕНЦИАЦИЯ ЧЕЛОВЕЧЕСТВАС момента возникновения Н. sapiens социальное в человеке стало его сущностью и биологическая эволюция видоизменялась, проявляясь в возникновении широкого генетического полиморфизма (см. § 12.3). Генетическое разнообразие на уровне генов и в меньшей степени хромосом обеспечивает разнообразие генотипов особей. Разнообразные генотипы по-разному проявляются в меняющихся условиях среды, давая огромное фенотипическое многообразие людей. В основе морфофизиологического полиморфизма человечества лежат полиморфизм наследственного материала на уровне генома и модификационная изменчивость. Эти факторы обеспечивают не только индивидуальное морфофизиологическое многообразие, но и внутривидовую групповую дифференциацию человечества на расы и адаптивные экологические типы. 15.4.1. Расы и расогенезНа протяжении длительного времени в антропологии господствовали представления о значимости расовой дифференцировки человечества и о большой роли естественного отбора в формировании основных расовых признаков. Применение методов молекулярной антропологии в значительной степени изменило представление о расах и расогенезе. Морфологические и в меньшей степени физиологические признаки дают возможность выделить внутри человечества три основные большие расы: европеидную, австрало-негроидную и монголоидную. Европеоиды имеют светлую или смуглую кожу, прямые или волнистые волосы, узкий выступающий нос, тонкие губы и развитый волосяной покров на лице и теле. У монголоидов кожа также может быть как светлой, так и темной, волосы обычно прямые, жесткие, темно пигментированные, косой разрез глаз и эпикант («третье веко»). Негроиды характеризуются темной кожей, курчавыми или волнистыми волосами, толстыми губами и широким, слегка выступающим носом. Имеются отличия рас и по некоторым физиологическим и биохимическим показателям: интенсивность потоотделения с единицы площади кожи у негроидов выше, чем у европеоидов, средние показатели уровня холестерина в плазме крови наиболее велики у европеоидов. В рамках каждой большой расы выделяются отдельные антропологические типы с устойчивыми комплексами признаков, называющиеся малыми расами. Существует три основных подхода к классификации рас: без учета их происхождения, с учетом происхождения и родства и на основе популяционной концепции. В соответствии с первым подходом три большие расы включают в себя 22 малые, причем между большими расами располагаются по две переходные малые. Схема расовой классификации изображается при этом в виде круга (рис. 15.7). Несмотря на то что при такой классификации не учитывается происхождение рас, само существование малых переходных рас, сочетающих в себе одновременно признаки двух больших рас (эфиопская, южносибирская, уральская и т.д.), свидетельствует, с одной стороны, о динамизме расовых комплексов признаков, а с другой — об условности членения человечества даже на большие расы. Гибридизация ДНК между большими выборками представителей малых рас в рамках одной большой показала высокую степень гомологии нуклеотидных последовательностей. Гибридизация ДНК представителей пар разных больших рас выявляет их значительную отдаленность друг от друга. Изучение гомологии нуклеотидных последовательностей западных европеоидов и представителей малой уральской расы и центрально-азиатских монголоидов с той же самой уральской расой дает среднее значение. Эти данные свидетельствуют о том, что переходные малые расы совмещают в себе не только морфологические признаки в соответствии с их промежуточным положением, но оказываются промежуточными и в отношении генетическом. Из этого следует, что они либо гибридогенны, либо сохранили в своей организации более древние черты, характерные для этапа существования человечества, предшествующего формированию больших рас. 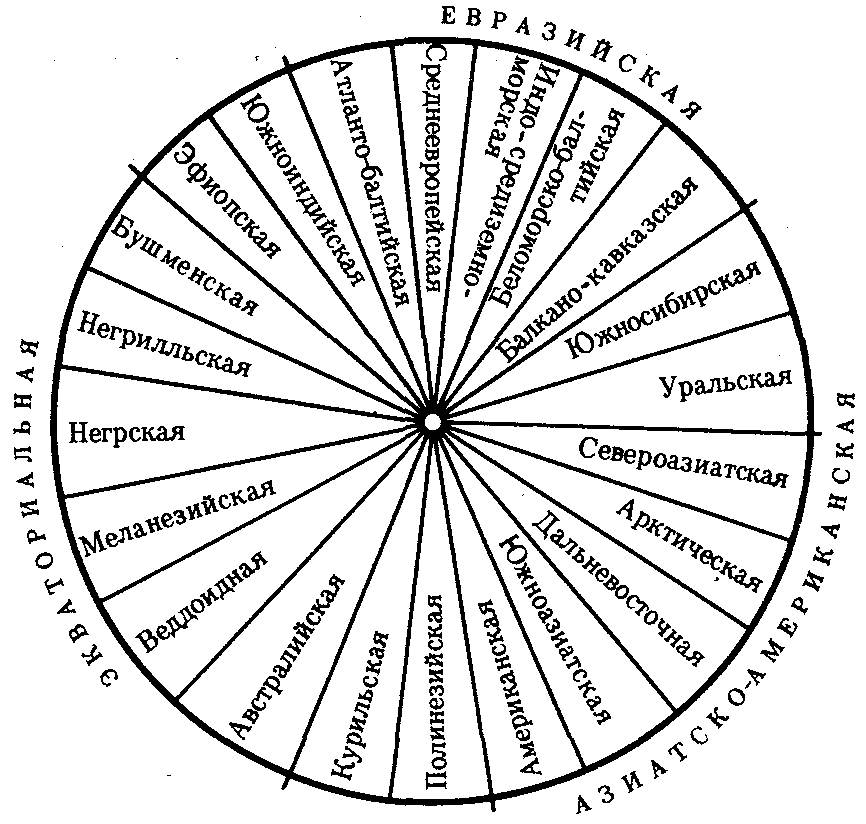 Рис. 15.7. Расовая классификация человечества без учета происхождения рас Классификация с учетом происхождения рас изображается в виде эволюционного древа с коротким общим стволом и расходящимися от него ветвями (рис. 15.8). В основе таких классификаций лежит обнаружение черт архаизма и эволюционной продвинутости отдельных рас, в соответствии с чем разные большие и малые расы занимают разное положение на ветвях такого древа. Выявление архаичных и прогрессивных черт среди морфологических признаков носит субъективный характер, благодаря чему схемы расовых классификаций такого рода очень многообразны. Но самым большим недостатком подхода к классификации рас исходя из их происхождения является попытка расположить расы на разных уровнях эволюционного древа, т.е. признание их биологической неравноценности. Кроме того, данные палеоантропологических исследований показывают, что вплоть до верхнего палеолита на территориях, обитаемых людьми, практически нигде не сформировались расовые типы человека, с которыми были бы генетически связаны современные большие расы. Это подтверждает анализ верхнепалеолитических находок скелетов людей современного физического типа из сунгирских погребений (Россия), живших приблизительно 26 тыс. лет назад. Все черепа, принадлежащие им, характеризуются мозаичным сочетанием расовых признаков и не могут быть отнесены ни к одной из современных рас. Этим данным соответствует и описание ископаемого скелета из Южной Калифорнии, пролежавшего в земле 21,5 тыс. лет и характеризующегося отсутствием выраженных монголоидных черт, несмотря на то что аборигенным населением Америки являются монголоиды. Только более поздние мезолитические находки свидетельствуют о формировании у человека расовых признаков. Так, известны мезолитические черепа с территории Северной Африки возрастом 10—8 тыс. лет с явными признаками не просто негроидной, а малой эфиопской расы. Сходные данные получены на территории Европы и в других регионах. Все это указывает на то, что процесс формирования расовых признаков — довольно поздний, идущий параллельно в разных регионах на рубеже верхнего палеолита — мезолита на фоне исходной разнородности расовых признаков у человека современного физического типа. Первичное появление на протяжении эволюции признаков малых, а не больших рас позволяет сделать вывод о том, что европеоидная, монголоидная и негроидная расы имеют мозаичное происхождение и представляют собой крупные популяции, объединенные не столько общностью происхождения, сколько климатогеографическими характеристиками условий существования и адаптивностью большинства основных признаков. 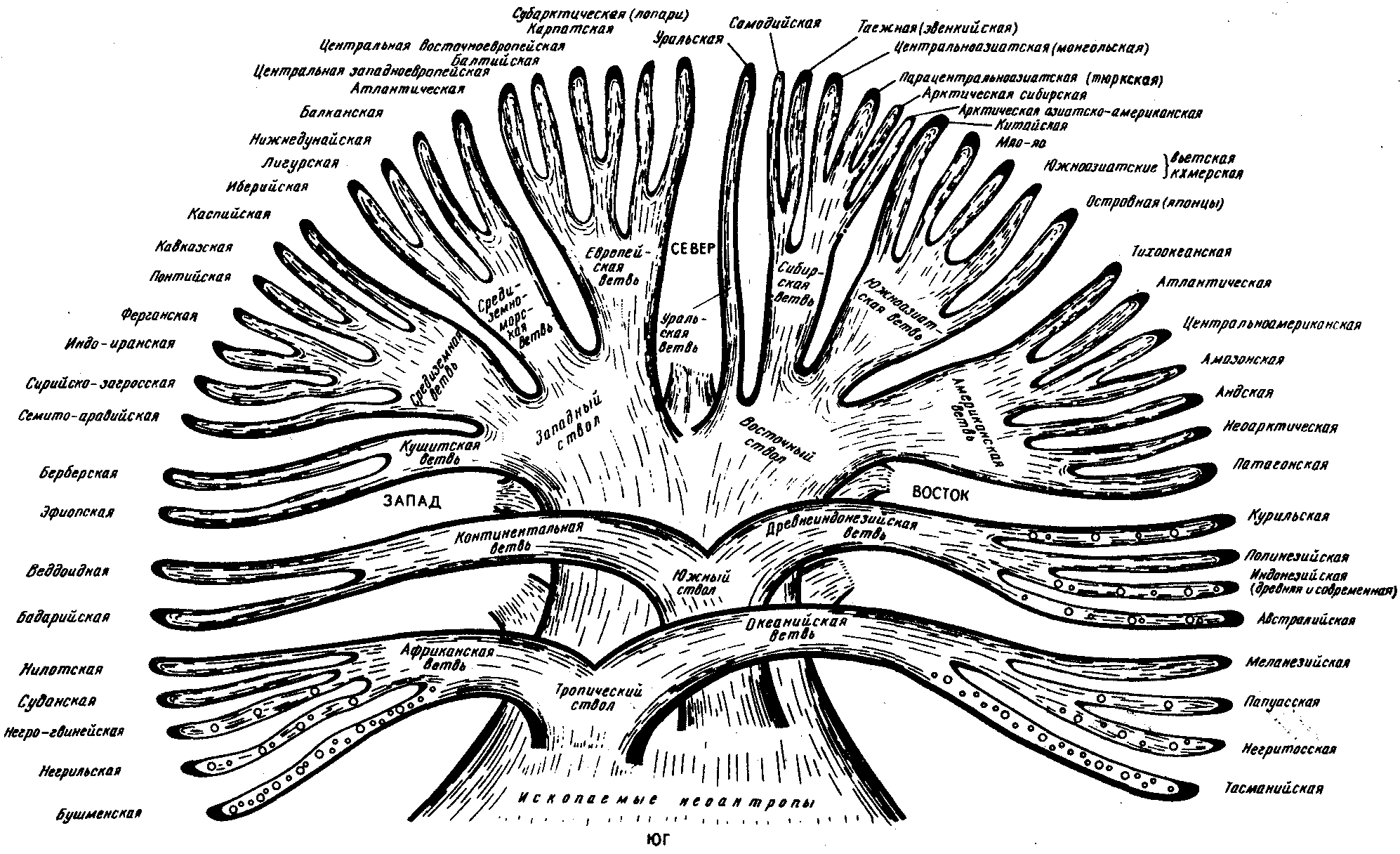 Рис. 15.8. Расовая классификация человечества с попыткой реконструкции происхождения рас Это заключение хорошо согласуется с популяционной концепцией рас. Суть ее заключается в следующем. Если принять, что большие расы человека представляют собой огромные популяции, то малые расы — субпопуляции больших, локальные естественные общности людей внутри которых — конкретные этнические образования (нации, народности) — являются более малыми популяциями. Если предположить при этом, намеренно упрощая ситуацию, что этносы не разделяются на элементарные популяции, и считать их просто состоящими из конкретных особей, то получится сложная структура, включающая в себя четыре уровня иерархии (рис. 15.9). 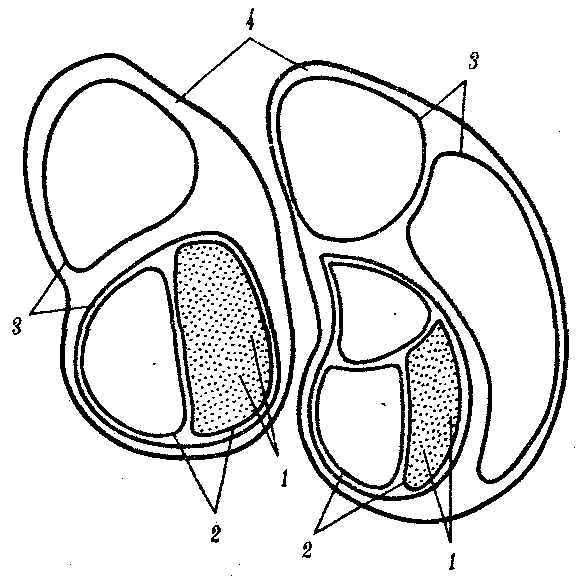 Рис. 15.9. Расы как выражение генетического полиморфизма человечества: 1—отдельные индивидуумы, 2—этносы, 3—малые расы, 4—большие расы На основании исследований распределения различных групп крови и белков в популяциях человека произведено сравнение доли каждого из четырех уровней меж- и внутрипопуляционных различий в общем объеме генетического полиморфизма человека по этим признакам.
Таким образом, от тотального генетического полиморфизма человечества расовые признаки составляют только 8%, в то время как основная доля генетического разнообразия определяется многообразием отдельных индивидуумов. Иными словами, немец может быть генетически гораздо ближе к полинезийцу, чем к другому немцу, живущему в соседней квартире. Изучение геногеографии популяций человека показало, что географическое распределение частот генов групп крови системы АВ0, MN, Lutheran, Duny, Diego и др., а также различных форм ферментов и иммуноглобулинов не соответствует ареалам расселения ни одной из рас. Так, по группам крови АВ0 и MN жители Европы оказываются ближе к африканцам, в то время как по системе иммуноглобулинов они ближе к монголоидам Азии. Сходные результаты получены и в отношении распределения в популяциях вариантов митохондриальной ДНК. Эти данные свидетельствуют о том, что биохимический полиморфизм человека эволюционно возник раньше и развивался дольше по сравнению с возникновением комплексов расовых признаков. Из этого следует, что расы не представляют собой особых изолированных групп людей, характеризующихся наборами специфических генов. Расовые же характеристики являются не более чем отдельными проявлениями общего генетического полиморфизма, выражающегося в первую очередь в сложных морфологических признаках. Некоторые из них адаптивны, другие сформировались на основе коррелятивной изменчивости, но все они касаются лишь ряда второстепенных особенностей (цвета кожи, волос, глаз и т.д.) и не затрагивают таких общечеловеческих признаков, как морфология головного мозга, а также строение и функции руки как органа труда. На основании определения числа аллелей, свойственных той или иной группе организмов, возможно определение генетического расстояния между ними. Эта величина для больших рас человека составляет 0,03. Она гораздо ниже цифр, характерных для истинных подвидов (0,17—0,22), и еще более мала по сравнению с межвидовым расстоянием (0,5—0,6 и более). В животном мире генетическое расстояние, равное 0,03, соответствует обычно генетическим отличиям местных популяций друг от друга. Все эти данные свидетельствуют о том, что понятие расы условно, второстепенно и не позволяет подводить под иерархическую классификацию рас глубокую биологическую, а значит, и социальную базу. 15.4.2. Адаптивные экологические типы человекаЧеловечество, заселившее уже около 15 тыс. лет назад все более или менее благоприятные для жизни природно-географические зоны, встретилось с необходимостью адаптироваться к самым разнообразным условиям существования. Адаптации человека к среде, как уже указывалось, проявляются в основном на социальном уровне, однако человечество на ранних этапах эволюции подвергалось непосредственному действию биотических и абиотических экологических факторов в значительно большей степени по сравнению с современной эрой научно-технического прогресса. Комплексы таких факторов имели разнонаправленное действие на человеческие популяции. В результате в разных климатогеографических зонах сформировались разнообразные адаптивные типы людей. Адаптивный тип представляет собой норму биологической реакции на комплекс условий окружающей среды и проявляется в развитии морфофункциональных, биохимических и иммунологических признаков, обеспечивающих оптимальную приспособленность к данным условиям обитания. В комплексы признаков адаптивных типов из разных географических зон входят общие и специфические элементы. К первым относят, например, показатели костно-мускульной массы тела, количество иммунных белков сыворотки крови человека. Такие элементы повышают общую сопротивляемость организма к неблагоприятным условиям среды. Специфические элементы отличаются разнообразием и тесно связаны с преобладающими условиями в данном месте обитания — гипоксией, жарким или холодным климатом. Именно их сочетание служит основанием к выделению адаптивных типов: арктического, тропического, зоны умеренного климата, высокогорного, пустынь и др. Разберем особенности условий жизни человеческих популяций в различных климатогеографических зонах и адаптивные типы людей, сформировавшиеся в них. Условия обитания в Арктике характеризуются постоянно низкими температурами воздуха, скудной растительностью, богатством животного мира и сезонной периодичностью поступления продуктов питания: растительная пища доступна только в короткие летние месяцы, а животная — в периоды нереста рыбы, гнездования птиц, размножения оленей и морского зверя. Продукты питания богаты витаминами, белками, жирами и микроэлементами, но бедны углеводами растительного происхождения. Воздух и почва в Заполярье содержат очень мало микроорганизмов, низкие температуры препятствуют сохранению цист патогенных простейших, яиц и личинок гельминтов. Из перечисленных факторов наибольшее влияние на формирование комплекса признаков арктического адаптивного типа оказали, по-видимому, холодный климат и преимущественно животная пища. Арктическому комплексу признаков свойственны относительно сильное развитие костно-мускульного компонента тела, большие размеры грудной клетки, высокий уровень гемоглобина, относительно большое пространство, занимаемое костным мозгом, повышенное содержание минеральных веществ в костях, высокое содержание в крови белков, холестерина, повышенная способность окислять жиры. Среди аборигенов Арктики почти не встречаются лица с астеническим телосложением. В целом арктический тип характеризуется усиленным энергетическим обменом, который отличается стабильностью показателей в условиях переохлаждения. Имеют свои особенности и механизмы терморегуляции. Так, при одинаковой степени охлаждения у канадских индейцев резко падает температура кожи, но уровень обмена веществ меняется незначительно, а у пришлого белого населения наблюдается меньшая степень снижения кожной температуры, но появляется сильная дрожь, т.е. интенсифицируется обмен. В тропиках и субтропиках располагается чуть ли не большая часть Ойкумены. Этот регион отличается в целом большим количеством тепла и влаги и сглаженностью сезонных колебаний условий обитания. Вместе с тем благодаря особенностям рельефа наблюдается значительная контрастность распределения тепла и влаги — массивы влажных лесов нередко соседствуют с засушливыми плато, обширными равнинами и редколесьем. В экваториальной и субэкваториальной областях сосредоточены огромные количества растительной биомассы. Влажные тропические леса относительно бедны животными, тогда как в саваннах животный мир разнообразен и включает крупных стадных животных, издавна используемых человеком как объект охоты. Почва, воздух и вода содержат большое количество микроорганизмов, яиц гельминтов и цист патогенных простейших. Богатство и разнообразие животного мира обеспечивает существование огромного количества промежуточных и окончательных хозяев биогельминтов и переносчиков возбудителей трансмиссивных заболеваний. К преобладающим экологическим факторам, под влиянием которых формировался комплекс признаков тропического адаптивного типа, относят жаркий влажный климат и рацион с относительно низким содержанием животного белка. В тропической области наблюдается исключительно широкая вариабельность групп населения в расовом, этническом и экономическом отношениях. Это проявляется в поразительном размахе изменчивости, например по соматическим признакам. Тем не менее преобладающие экологические факторы, особенно климатический, способствовали образованию определенного комплекса морфофизиологических признаков обитателей тропиков и субтропиков. К характерным признакам тропического типа относят удлиненную форму тела, сниженную мышечную массу, относительное уменьшение массы тела при увеличении длины конечностей, уменьшение окружности грудной клетки, более интенсивное потоотделение за счет повышенного количества потовых желез на 1 см2 кожи, низкие показатели основного обмена и синтеза жиров, сниженную концентрацию холестерина в крови. Антропологическое изучение современных обитателей зоны умеренного климата под углом зрения формирования биологических механизмов адаптации к природным условиям затруднено, так как значительная часть людей проживает в промышленно развитых странах с большой долей городского населения. Результаты наблюдения позволяют, однако, судить о том, что и в этом случае в процессе исторического развития человеческих популяций сформировался комплекс признаков, соответствующий особому адаптивному типу умеренного пояса. По соматическим показателям, уровню основного обмена население умеренного пояса занимает промежуточное положение между коренными жителями арктического и тропического регионов. Это соответствует условиям биогеографической среды в зоне умеренного климата. Для нее характерны неравномерное распределение районов, отличающихся по количеству тепла и влаги, типу растительности (от сухих степей и полупустынь до тайги), богатству животного мира. Вместе с тем температура и влажность воздуха здесь не достигают экстремальных величин, хорошо выражен сезонный ритм биоклиматических условий. Условия высокогорья для человека во многих отношениях экстремальны. Их характеризуют низкое атмосферное давление, сниженное парциальное давление кислорода, холод, относительное однообразие пищи. Основным экологическим фактором формирования горного адаптивного типа явилась, по-видимому, гипоксия. У жителей высокогорья независимо от климатической зоны, расовой и этнической принадлежности наблюдаются повышенный уровень основного обмена, относительное удлинение длинных трубчатых костей скелета, расширение грудной клетки, повышение кислородной емкости крови за счет увеличения количества эритроцитов, содержания гемоглобина и относительной легкости его перехода в оксигемоглобин. 15.4.3. Происхождение адаптивных экологических типовЧеловечество возникло в тропической зоне Африки, и следует полагать, что поэтому наиболее древним и исходным для остальных экологических типов человека является тропический тип. Об этом свидетельствует и наибольший полиморфизм морфофункциональных признаков человека в рамках этого типа на территории Центральной и Северо-Восточной Африки. В связи с этим уместно обратиться к учению Н.И. Вавилова о центрах происхождения и многообразия культурных растений, в соответствии с которым именно в зонах первоначального вовлечения биологических видов в социальную среду наблюдается наиболее выраженный наследственный полиморфизм местных популяций по большому количеству признаков. Об этом свидетельствует и то, что именно в тропической Африке в непосредственной близости друг от друга, т.е. под действием сходных экологических факторов, обитают наиболее низкорослые и наиболее высокорослые племена, например пигмеи, готтентоты и бушмены, с одной стороны, и массаи — с другой. Пределы изменчивости африканских популяций, например, по росту и массе тела таковы, что в них укладываются все известные на Земле человеческие популяции. Основные же черты тропического типа в Африке, отмеченные выше, остаются неизменными. Они же характеризуют монголоидные популяции Индокитая, Малайского архипелага и некоторые группы индейских племен зоны влажных тропических лесов Центральной и Южной Америки. Это указывает на то, что тропический адаптивный тип развивается в результате асинхронного параллелизма в эволюции человека, причем вначале на африканском континенте, а потом и в других областях (см. раздел 3.13). Отсюда следует и еще один вывод: адаптивный тип формируется на фоне расогенеза и вне зависимости от него. В процессе адаптогенеза в популяциях человека можно проследить и проявление закона гомологических рядов (см. разд. 13.3.5). Это выражается, например, в том, что в бассейне Меконга в Индокитае и на острове Суматра имеются пигмеоидные популяции, по антропометрическим признакам соответствующие африканским пигмеям. Экологический тип умеренного пояса сформировался на базе исходного генетического и фенотипического полиморфизма тропического типа при расселении популяций человека в умеренных зонах Евразии и позже — Северной Америки. Он оформился в рамках двух больших рас: европеоидной и монголоидной. При заселении человеком арктической зоны в Евразии и Северной Америке произошло формирование арктического типа. Независимо от него среди индейцев Южной Патагонии и Огненной Земли в приантарктической зоне Южной Америки возникли популяции индейцев, по основному комплексу признаков соответствующие арктическому типу. Это еще один убедительный пример параллелизма эволюции человеческих популяций и реализации закона гомологических рядов, а также доказательство вторичности адаптивных типов по отношению к большим расам человечества. Формирование горного адаптивного типа иллюстрирует общие закономерности адаптогенеза, отмеченные выше. Этот тип также развился независимо от расовой и этнической принадлежности популяций — среди европеоидов Альп, Кавказа, Памира и Гималаев, а также в монголоидных популяциях Тибета, Тянь-Шаня и Анд. В связи с тем что высокогорья заселялись человеком в последнюю очередь, горный экологический тип является по происхождению самым молодым. Интересно, что, несмотря на особенно выраженную расовую и этническую разнородность этого типа, комплекс основных признаков его является монолитным. Однако в отличие от других типов, вероятно, именно горный проявляется в основном только на фенотипическом уровне и не имеет наследственной природы. Об этом свидетельствует то, что количество эритроцитов в крови и объем грудной клетки людей, переселяющихся в условия высокогорья и обратно, могут меняться на протяжении жизни одного поколения. Следовательно, адаптивные типы человека не только отражают его прошлое, но формируются и в настоящем, а разные типы имеют и разную по длительности историю. В настоящее время пока еще на фенотипическом уровне идет формирование адаптивного типа человека городской среды, который характеризуется широкой лабильностью психических реакций, обеспечивающих способность переживать состояния постоянного стресса, и рядом морфофизиологических особенностей, оптимальных для жизни в специфических условиях города (см. разд. 17.3.2). Формирование экологических типов человека в значительной степени обеспечило всесветное расселение людей. Меняющаяся среда обитания под действием антропогенных факторов ставит перед популяциями человека новые задачи, решаемые и сегодня за счет не только социальных, но и биологических адаптации. Схему эволюционных взаимоотношений больших рас и адаптивных типов человека см. на рис. 15.10. 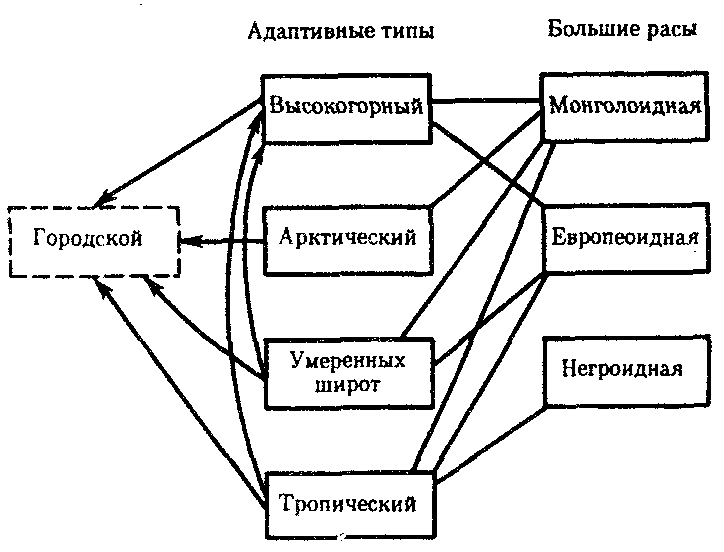 Рис. 15.10. Адаптивные типы человека и большие расы |