Обмен липидов.
Липиды:
1). Распад 3-ацилглицерина (ТАГ) в ткани катализирует липаза (гормончувствительная).
Механизм активации тканевой липазы
Гормон (+глюкагон,-инсулин)
↓
н-аденилатциклаза → а-аденилатциклаза

 АТФ → цАМФ + Н4P2O7 АТФ → цАМФ + Н4P2O7
н-ПК-А → а-ПК-А
 н-тканевая липаза → а-тканевая липаза н-тканевая липаза → а-тканевая липаза
АТФ АДФ
ТАГ → глицерин + жирные кислоты
+Н2О
2). Окисление глицерина: глицеролфосфат-
Глицеринкиназа дегидрогеназа
Г    лицерин а-глицеролфосфат лицерин а-глицеролфосфат
АТФ АДФ НАД+ НАДН2
дигидроксиацетонфосфат → далее идёт гликолиз
3).окисление жирных кислот:
начинается с активации жирной кислоты(R-COOH)
ацил-КоАсинтетаза
R -COOH + KoA-SH + ATФ ацил-КоА + АМФ +Н4P2O7 (активированная -COOH + KoA-SH + ATФ ацил-КоА + АМФ +Н4P2O7 (активированная
жирная кислота)
Эта реакция протекает в цитоплазме,а собственно окисление жирных кислот в матриксе митохондрий. Короткоцепочечные молекулы проходят самостоятельно, а длинноцепочечные с помощью карнитина.

цитозоль внутренняя мембрана матрикс

  ацил-КоА карнитин ацилКоА ацил-КоА карнитин ацилКоА
 
1 2
КоА ацилкарнитин КоА
1 и 2- карнитинацилтрансфераза. Жирные кислоты вступают в в-окисление – цикл Кноопа-Линена:
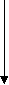 Ацил-КоА Ацил-КоА
ФАД
АцилКоА-дегидрогеназа
ФАДН2
еноил КоА
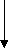
+Н2О еноилКоА-гидротаза
-гидроксиацилКоА
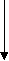  НАД+ НАД+
НАДН2 -гидроксиацилКоА-дегидрогеназа
-кетоацилкоА
HS-KoA→ тиолаза
Ацил-КоА + Ацетил-КоА
На этом заканчивается первый виток цикла. В ходе этого витка образуется 1 ФАДН2, 1НАДН2 и ацетил КоА→ цикл Кребса
Образовавшийся новый ацилКоА в следующие аналогичные витки цикла. Последний виток вступает в бутирилКоА, который распадается на 2 ацетилКоА. В последнем витке образуется такое же количество коферментов.
Расчёт энергетической ценности жирных кислот:
С16 : 1). 8 ацетилКоА (16:8=2)
2). количество витков: 8-1=7
3). АТФ: 7НАДН2- 21 АТФ
7ФАДН2- 14АТФ
4). на активацию жирной кислоты 1 АТФ
Всего АТФ 21+14-1=34
12*9
2*8
3*8
-1
Итог: 147 АТФ
Если у жирной кислоты появляется двойная связь, то не будет ФАД и не будет первой реакции
Особенности окисления жирных кислот с нечётным числом углеродных атомов заключается в том, что последний виток вступает к 5 углеродным фрагментам, ацетилКоА, пропионилКоА – не является в цикле Кребса, поэтому пропионилКоА должен превратиться в один из метаболитов цикла Кребса.
ПропионилКоА
 СО2 АТФ СО2 АТФ
АДФ + Н3РО4 пропионилКоАкарбоксилаза
МетилмалонилКоА
метилмалонилКоАмутаза
сукцинилКоА → цикл Кребса
4).Синтез липидов : синтез жирных кислот протекает в ЭПС разных тканей на полиферментном комплексе, который называется синтаза жирных кислот или пальмитатсинтаза.
Комплекс состоит из 7 ферментов, в центре находится ацилпереносящий белок (АПБ), а вокруг него 6 остальных ферментов. Комплекс имеет свободные SH-группы, принадлежащие 4-фосфопантатеину. На периферии это остатки цистеина. Синтез жирных кислот начинается с ацетилаКоА, который поступает в цитоплазму из матрикса митохондрий в составе субстрата.
Цитрат + КоА-SH + АТФ → ацетилКоА + оксалоацетат + АДФ + фосфорная кислота; фермент- АТФцитратлиаза.
Из ацетилКоА образуется малонилКоА, который является основным субстратом для синтеза жирных кислот.
АцетилКоА
  АТФ АТФ
 СО2 ацетилКоАкарбоксилаза СО2 ацетилКоАкарбоксилаза
АДФ +Н3РО4
малонилКоА
1).перенос ацетила с ацетилКоА на синтетазу; фермент- ацетилтрансацилаза.
2).перенос малонила с малонилКоА на синтетазу; фермент- малонилтрансацилаза.
3).конденсация ацетила с малонилом и декарбоксилирование образовавшегося продукта; фермент- в-кетоацил-синтетаза
4).первое восстановление промежуточного продукта с участием НАДФН2; фермент- в-кетоацил-редуктаза.
5).дегидратация промежуточного продукта; фермент- гидроксиацил-гидратаза
6).второе восстановление промежуточного продукта с участием НАДФН2; фермент- еноилредуктаза.
Образовавшийся бутирильный остаток переносится на верхнюю SH-группу с помощью ацетилацилтрансферазы, а затем образуется новый остаток и синтез повторяется до тех пор, пока не образуется кислота нужной длины.
Синтез кислот регулируется гормонами:
1).активность ацетилКоА-карбоксилазы стимулируется инсулином, тормозится глюкагоном и длинноцепочечными жирными кислотами.
2).синтаза жирных кислот активируется инсулином, тормозится глюкагоном
Синтез ТАГ протекает в цитоплазме разных клеток
-глицеролфосфат- фосфати-
ацилтрасфераза датфосфатаза
а  -глицеролфосфат фосфатидная кислота -глицеролфосфат фосфатидная кислота
ДАГацилтрансфераза Н2О Н3РО 4
д иацилглицерин триацилглицерин иацилглицерин триацилглицерин
Схема превращения глюкозы в ТАГ
глюкоза
 АТФ АТФ
АДФ
Глюкозо-6-фосфат → ПФЦ
Фруктозо-1,6-бисфосфат
  дегидроксобисфосфат дегидроксобисфосфат
Глицеральдегид-3-фосфат -глицеролфосфат НАДН2
НАД+
ПВК ТАГ
ацетилКоА → ацилКоА
НАДФН2 |
 Скачать 49.5 Kb.
Скачать 49.5 Kb.