Рибосома СӨЖ. Рибосомалар. Негізгі блім Рибосоманы ашылу тарихы
 Скачать 0.53 Mb. Скачать 0.53 Mb.
|

Жоспар: Кіріспе Негізгі бөлім Рибосоманың ашылу тарихы Прокариот және эукариот рибосомаларының айырмашылығы. Рибосома структурасы Рибосоманың белок биосинтезіндегі рөлі. Қорытынды Пайдаланған әдебиет Кіріспе Рибосомалар зерттеу жүргізілген өсімдік клеткаларының барлығынан табылған. Рибосомалардың болатынын алғаш рет 1941 жылы Касперсон мен Браше атап көрсетті. Рибосомаларға сипаттама беру үшін седиментация коэффициентін қолданады, оны ерітіндідегі бөлшектердің тұну жылдамдығы бойынша анықтайды. 'Бұл коэффициент рибосомалардың өлшемдері мен салмағына байланысты. Бұл шаманы 8 таңбасымен белгілейді, яғни ол Сведберг бірлігін білдіреді. А. С. Спирин. мен Д. П. Гаврилова (1968) прокариоттардан алынған рибосомалардың седиментация коэффициенті 708, ал эукариоттардан алынған рибосомалардыц седиментация коэффициенті 808 болуын анықтады. Рибосомалар - бұл барлық тірі жасушаларда кездесетін, биологиялық ақуыз синтезінің (мРНҚ) трансляциясын жүзеге асыратын жоғары молекулалық машиналар. Рибосомалар аминоқышқылдарды кодондарда көрсетілген ретпен байланыстырады, олар матрицалық РНҚ (мРНҚ) молекулаларынан полипептидтік тізбектер түзеді. Рибосома тірі организмдердің барлығынан дерлік табылған. Әр жасушада шамамен 1000-нан 10000-ға дейін Рибосомалар болады. Шамамен диам. 20 нм-дей. Рибосомалар екі класқа бөлінеді: 70S және 80S (S-седиментация коэфф., Сведберг бірлігі). 70S Рибосома ядросы жоқ прокариоттар жасушаларында, ал 80S Р-лар эукариоттардың цитоплазмасында болады. Химиялық құрамы жағынан Р-лар РНІ-нан және ақуыздардан тұратын нуклеопротеид болып саналады. 70SBold text Рибосомасының 60 — 65%-ы РНҚжәне 35 — 40%-ы ақуыздан, ал 80S Рибосомасының 50%-ы РНҚ-дан, 50%-ы ақуыздан құралады. Құрылымы жағынан Рибосомалар үлкен және кіші суббөліктерден тұрады. Рибосомалар цитоплазмада топтасып (5-70-тен) орналасып, полисомалар (полирибосома) түзеді. Эукариотты жасушаларда рибосомалардың көп бөлігі ЭПТ-ға жабысады, нәтижесінде ол кедір-бұдыр болады. Эукариоттық жасушаның рибосомаларының құрамына нуклеотидтер санымен ерекшеленетін рибосомалық РНҚ (ррнқ) төрт түрі кіреді: 18s-1900 нуклеотидтері; 5S-120 нуклеотид; 5,8 s-160 нуклеотид; 28s-4800 нуклеотид. 18s-ррнқ және 30-35 ақуыз кіші бөлімді құрайды, қалған рибонуклеин қышқылдары мен 45-50 ақуыз үлкен субунит құрамына кіреді. Ядрода ішкі бөліктер бөлек синтезделеді. Олар бір рибосомада тек жұмыс үшін жиналады – матрицалық РНҚ-да болатын ақуыз синтезі. Бөлімшелер mRNA-ны орап, полисомалар немесе полирибосомалар деп аталатын кешендерге жиналады. біріктіріледі.Құрылымы бойынша рибосоманың екі бөлімі де белгілі бір үшіншілік құрылымдарды қабылдайтын (бүктелген) және ондаған түрлі ақуыздармен қапталған РНҚ молекулалары болып табылады. Бұл жағдайда рибосомалардың үлкен бөлігінің құрамына үш рРНҚ молекуласы кіреді (прокариоттарда — екеуі), ал кіші бөліктің құрамына тек біреуі кіреді Прокариоттардың үлкен бөлігіне РНҚ-ның екі түрі, ал кішісіне біреуі кіредіПрокариоттардың рибосомалары ұқсас құрылымға ие, бірақ олар эукариоттықтардан біршама кіші (толық рибосоманың шөгу коэффициенттері 70S, ал суббірліктердің 30S және 50S). Митохондрия мен хлоропластардың рибосомалары прокариотқа жақын. Прокариоттық және эукариотты рибосомалар - тірі жасушалардың құрамдас бөлігі.Олар ақуыз синтезі үшін сайт ұсынады. Сонымен қатар, олар РНҚ мен рибосомалық ақуыздардан жасалады. Сондай-ақ, олардың құрамында РНҚ-дан тұратын екі бөлім бар. Сонымен қатар, екеуі де жасуша цитоплазмасында болады.Сонымен қатар, олардың кіші бөлімі РНҚ-ның бір тізбегінен тұрды.
Құрылымы бойынша рибосомалар кішкентай, дөңгелек пішінді, мембранасыз жасуша органоидтары. Олардың әртүрлі жасушалардағы саны мыңнан бірнеше миллионға дейін өзгереді. Рибосома монолитті құрылым емес, ол үлкен және кіші бөліктер деп аталатын екі бөлшектен тұрады. Клиникада кеңінен қолданылатын тетроциклиндер 70S рибосомасында нәруыздың синтезделуіне ингибитор болып табылған(80S рибосомадағы синтез аз тежеледі). Олар жасушалық мембраналар арқылы жеңіл өтеді. Тетрациклиндер 50S рибосома суббөлшегіндегі т-РНҚ аминоацилдің аминоацилді орталықпен байланысуын тежейді деп есептеледі. Тетрациклиндер трансляция процесінің алдыңғы кезеңдерінің бірін қоса, осы орталықпен химиялық байланысуы мүмкін. Матрицалық РНҚ, транспорттық РНҚ, рибосомадағы көптеген ақуыз факторлары белгілі бір позицияларды алады, бұл химиялық реакциялардың тиімді өтуіне мүмкіндік береді. Рибосомалар екі негізгі компоненттен тұрады   Рибосомадағы суббірліктерді біріктірген кезде – сайттар пайда болады. Рибосома мРНҚ арқылы қозғалады және кодондарды бірінен кеиін бірін "оқиды". Бір сайтқа оған қосылған амин қышқылы бар тРНҚ кіреді, екіншісінде – бұрын синтезделген полипептидтік тізбек бекітілген тРНҚ бар. Рибосомада амин қышқылы мен полипептид арасында пептидтік байланыс пайда болады. Нәтижесінде полипептид "жаңа" тРНҚ-ға түседі, ал "ескі" рибосомадан кетеді. Қалған тРНҚ оның орнына "құйрығымен" (полипептид) ауысады. Рибосома мРНҚ арқылы бір триплетке алға жылжиды және оған қосымша РНҚ қосылады. Рибосомалар екі түрлі қосалқы суббөлшектерден тұрады, олардың әрқайсысы рибосомалық РНҚ [ррнқ (rRNA)] және көптеген ақуыздардан тұрады. Рибосомалар мен олардың қосалқы бөлшектері әдетте масса бойынша емес, тұндыру коэффициенттеріне сәйкес жіктеледі. Сонымен, толық эукариоттық рибосоманың тұндыру коэффициенті шамамен 80 Сведберг (80S), ал оның қосалқы бөлшектерінің тұндыру коэффициенті 40S және 60s құрайды. Рибосомалардың орналасуына байланысты екі түрі бар: байланысқан-ЭПС-ке қоныстану; бос-цитоплазмада болады. Бөлімшелер екі түрге бөлінеді – үлкен және кіші. Әр бөлік нуклеин қышқылдары мен ақуыз қоспасынан тұрады, яғни рибосома химиялық құрылымы бойынша нуклеопротеид болып табылады.  Рибосома құрылысы: 1 – кіші суббірлік; 2 – аРНҚ; 3 – тРНҚ; 4 – аминқышқылы; 5 – үлкен суббірлік; 6 – эндоплазмалық тордың мембранасы; 7 – синтездеуші полипептидті тізбек.        Белок синтезі - өте күрделі процесс. Белок синтезінің негізінде жатқан молекулалық процестер өте күрделі. Олардың көпшілігі жазылып суреттелгенмен толық мазмұнын, айталық транскрипция, репарация және ДНҚ-ның репликациясы тәрізді түсіндіру әзір мүмкін емес. Мысалы, белок синтезінде РНҚ молекулаларының кез келген бір класы емес, үш класы (аРНҚ, тРНҚ және рРНҚ) қатысады, бірақ неге бұлай болатыны айқын түсінікті емес. Сондықтан белок синтезінің егжей-тегжей негізінен, әліде белгілі бір теорияда жалпыланбаған жалаң факты ретінде қабылдауымыз керек. Белок синтезінің процесінде басты агент ролін тРНҚ молекулалары атқарады. Оларға полимеризацияланбай тұрып, яғни полипептидтерге бірікпей тұрып, амин қышқылдары жалғасады. тРНҚ-ның молекуласына карбоксилдік ұшымен қосыла отырып, амин қышқылдары белсенді түрде энергияға бай түрге айнала ды, ол өз бетімен пептидтік байланыс түзе алады, сөйтіп полипептидтерді синтездеуге мүмкіндік туады. Бұл белсенділік процесі - белок синтезіне қажетті кезең, себебі бос амин қышқылдары полипептидтік тізбекке тікелей жалғаса алмайды. Өсіп келе жатқан полипептидтік тізбекке дәл сол амин қышқылы қосылуы керектігі амин қышқылына байланысты емес, оны тіркеп алған тРНҚ молекуласына тәуелді. Мұны бір ерекше әсем тәжірибенің көмегімен анықтауға мүмкін болды, онда ерекше тРНҚ-ға жалғанған амин қышқылын химиялық әдіспен басқа амин қышқылына (цистеинді аланинге) айналдырған. Кейін мұндай будан молекулалар клеткасыз жүйеде жұмыс істегенде, дұрыс емес амин қышқылы белок тізбегіне сол тРНҚ "қызмет" жасағанда үнемі қосылып отырған. Белок биосинтезіне қатысатындар:    Аминқышқылдар Ферменттер Рибосомалар РНҚ Белоктардың синтезі бір рибосомада өтуі мүмкін немесе бір уақытта бірнеше рибосомада не полисомада/ жүруі мүмкін. Полисома бір и-РНҚ бойында бола алатын рибосомалар тобы /80-ге жуық рибосома/ болуы мүмкін. Мұндай бір и-РНҚ-ның бойындағы информацияны бір уақытта бірнеше рибосоманың көмегімен белок синтезіне қолдану синтездің тез және тиімді өтуіне мүмкіндік тудырады. Бактерияларда транскрипция және трансляция бірімен-бірі ілесіп жүреді, яғни ДНҚ-на тәуелді РНҚ-полимераза и-РНҚ-ның синтезін жүргізіп жатқан кезде, и-РНҚ-ның бір шетінде белок синтезі де басталып жатады. Бактериялардың екінші бір ерекшелігі и-РНҚ-ның тіршілік ету уақыты бірнеше минут қана, сонан соң олар тез нуклеаза ферментінің әсерімен ыдырап кетеді. Белоктардың синтезі көптеген антибиотиктер әсерінен тежеуге ұшырауы мүмкін. Кейбір микроорганизмдер үшін қорғаныш антибиотиктер, басқа организмдер үшін өте улы болып табылады. Мысалы: пурамицин - элонгация кезеңінде әсер етсе, тетрациклин аминоацил - т-РНҚ-ның рибосомадағы аминоацилдік центрімен байланысуына кедергі жасайды; стрептомицин - рибосоманың кіші суббірлігімен қосылып оның қызметін нашарлатады; дифтерия токсині-элонгация факторын тежейді; левомицетин - пептидилтрансфераза ферментінің активтілігін нашарлатады; эритромицин - үлкен суббірлікпен қосылып, транслоказа ферментінің жұмысын тежейді. Белоктар синтезінің реттелуі. Белок синтезінің реттелуі и-РНҚ-ның синтезі және трансляция /яғни белок синтезі/ кезеңінде жүреді. Бұл бағытта аса көп жұмыс істеген француз ғалымдары Жакоб және Моно болды. Бұл ғалымдар осы жұмысы үшін Нобель сыйлығына ие болды. Олар белоктарды синтездеу теориясын оперон теориясы деп атады. Бұл ғалымдардың пікірі бойынша бактерияларда ең кемінде геннің үш түрі болады: I/ оператор гені /0-ген/; 2/ реттеуші ген / R -ген/; 3/ белоктардың бірінші реттік құрылысын анықтайтын құрылымдық ген / S - ген/. ДНҚ молекуласының осы үш ген орналасқан бөлімін оперон деп атайды да, бірімен-бірі тығыз байланысты болады. Реттеуші ген оператор геніне репрессор арқылы әсер етіп отырса, оператор гені құрылымдық генге әсер етеді Барлық ферменттік белоктардың синтезін реттеуді үш топқа бөлуге болады: репрессибилді, яғни белоктардың синтезін тежеу; индуцибелді, белок синтезінің жылдамдығын арттыру; конституитивті немесе кейбір белоктар синтезінің жылдамдығының тұрақты болуы. 1.Белоктардың синтезін тежеу немесе репресибилді жүйелер кебінесе анаболизм реакцияларына қатысатын ферменттердің синтезінде қолданылады. Мұндай жүйелерде құрылымдық гендер / S - гендер/ тұрақты жұмыс істеп тұрады. Реттеуші геннің қатысуымен активсіз белок - репрессор синтезделеді. Енді осы белок - репрессорды активті күйге көшіру үшін корепрессор қажет. Корепресеордың қызметін кейбір кіші молекулалы заттар, реакция нәтижесінде түзілген немесе реакция аралық заттар, гормондар атқара алады. 2. Белок синтезінің жылдамдығын арттыру немесе индуцибелді жүйелер. Бұл жүйе түрінде реттеу катаболизм реакцияларына тән. Мұндай жүйелерде құрылымдық гендер сыртқы орта туғызған жағдайларға тәуелді, яғни клеткаға катаболизм реакцияларына қатысатын ферменттер қажет болғанда ғана жұмыс істейді. Бұл жүйелерде реттеуші геннің қатысуымен синтезделетін белок - репрессор активті болады да, ол оператор генімен комплекс түзеді. Сондықтан оператор гені құрылымдық гендердің жұмысын қамтамасыз етпейді. Бірақ активті белок - репрессордың сыртқы ортадағы клеткаға түскен төменгі молекулалы заттармен қосылыс түзіп активсіз күйге көшетін қасиеті бар. Ол кезде оператор гені белок-репрессордан босап, құрылымдық гендердің жұмысы басталады, яғни и-РНҚ құрылымдық гендердегі сол клеткаға түскен заттардың катаболизмін қамтамасыз ететін ферменттердің бірінші реттік құрылысын жазып алады. Белок синтезінің осы индуцибелді жолмен реттелуі Е. colire жүргізілген тәжірибелер арқылы дәлелденген. Е. coli әдетте тек глюкозамен ғана қоректенеді. Ал егер осы ортаға лактозаны қоссақ, оны галактоза мен глюкозаға ыдырататын лактаза / β - галактоэидаза/ ферменті синтезделгенше, микроорганизмдердің өсуі біраз уақытқа тоқтайды. Клеткаға түскен лактоза /индуктор қызметін атқарады/ активті белок репрессормен қосылып оператор генінің балок - репрессормен қосылуына кедергі жасайды. Соның арқасында оператор гені мен құрылымдық гендер қажетті и-РНҚ түзілуіне, ал ол рибосомада лактозаны ыдыратуға қажет β -галактозидаза ферментінің синтезін қамтамасыз етеді. 3. Конституитивті немесе синтезделу жылдамдықтары тұрақты болатын белоктар. Мұндай белок - ферменттерінің құрьшымдық гендері тұрақты жұмыс істейді де, басқа геңдердің ықпалы әсер етпейді. Бұл ферменттердің қатарына гликолиз, үш карбон қышқылдарының цикліне қатысатын ферменттер жатады. 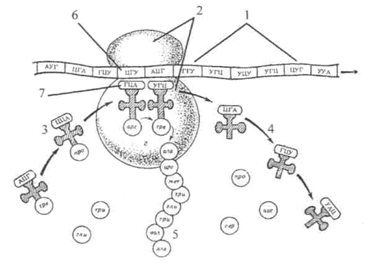 Белок биосинтезі: 1 — и-РНК; 2 – рибосоманың субъединицасы; 3 — т-РНК аминкышқылымен; 4 — т-РНК аминкышқылсыз; 5 — полипептид; 6 — кодон а-РНК; 7- антикодон т-РНҚ Белок биосинтезі: 1 — и-РНК; 2 – рибосоманың субъединицасы; 3 — т-РНК аминкышқылымен; 4 — т-РНК аминкышқылсыз; 5 — полипептид; 6 — кодон а-РНК; 7- антикодон т-РНҚҚорытынды Рибосомалар - бұл барлық тірі жасушаларда кездесетін, биологиялық ақуыз синтезінің (мРНҚ) трансляциясын жүзеге асыратын жоғары молекулалық машиналар. Рибосомалар аминоқышқылдарды кодондарда көрсетілген ретпен байланыстырады, олар матрицалық РНҚ (мРНҚ) молекулаларынан полипептидтік тізбектер түзеді. Әрбір бөлімше бір немесе бірнеше рибосомалық РНҚ (рРНҚ) молекулаларынан және көптеген рибосомалық ақуыздардан (RP немесе r-ақуыздар) тұрады Рибосомаларды алғаш рет 1950 жылдардың ортасында румын-америкалық жасуша биологы Джордж Эмиль Паладе электронды микроскопты пайдалана отырып тығыз бөлшектер немесе түйіршіктер түрінде ашқан. Ричард Робертс 1958 жылы «микросомалық фракцияның рибонуклеопротеиндік бөлшегін» «Рибосома» терминіне ауыстыруды ұсынды. Альберт Клод, Кристиан де Дюв және Джордж Эмиль Паладе бірге физиология немесе медицина саласындағы Нобель сыйлығын 1974 жылы рибосоманы тапқаны үшін алды. Рибосома - өте күрделі жасушалық аппарат. Ол көбіне рибосомалық РНҚ (рРНҚ) деп аталатын мамандандырылған РНҚ-дан, сондай-ақ ондаған әртүрлі ақуыздардан тұрады (түрлер саны арасында аз мөлшерде өзгереді). Рибосомалық ақуыздар мен рРНҚ әр түрлі көлемдегі екі бөлек рибосомалық бөліктерге бөлінген, оларды әдетте үлкен және кіші рибосомалық суббірліктер деп атайды. Пайдаланған әдебиет С. Ж. Стамбеков, В. Л. Петухов. Молекулалық биология. Новосибирск-2003г. А. Ж. Сейтембетова, С. С. Лиходий. Биологиялық химия. Шаффер, Екатерина. «Рибосоманың құрылымы.» News-Medical.net, News Medical, 2018ж https://obrazovaka.ru/biologiya/stroenie-ribosomy-osobennosti.html |