био вопросы 4 тема. Открытие днк и доказательства ее генетической роли
 Скачать 0.49 Mb. Скачать 0.49 Mb.
|

|
9. Понятие о гене. Свойства гена. Ген – участок молекулы ДНК, кодирующий последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекуле тРНК или рРНК. Свойства гена: Дискретность — несмешиваемость генов; Стабильность — способность сохранять структуру; Лабильность — способность многократно мутировать; Множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм; Аллельность — в генотипе диплоидных организмов только две формы гена; Специфичность — каждый ген кодирует свой признак; Плейотропия — множественный эффект гена; Экспрессивность — степень выраженности гена в признаке; Пенетрантность — частота проявления гена в фенотипе; Амплификация — увеличение количества копий гена. В 1902 году английский врач Гаррод описал насле-дственную болезнь алкаптонурию , обусловленную нарушением метаболизма гомогентезиновой кислоты. Это свидетельствует о том, что гены контролируют определенные метаболические функции. Это подтвердили Бидл и Тэйтум в1940 году опытами с мутантами хлебной плесени Neurospora crassa, вы-двинув гипотезу – «один ген – один фермент». Полученные позже данные о химическом составе и строении ДНК позволили сформулировать концепцию о том, что генетическая информация в полинуклеотидной цепи ДНК определяется порядком чередования в ней нуклеотидов. В 1961 году Ф.Крик и С.Бензер показали, что кодирование аминокислот в молекулах белка осуществляется триплетом нуклеотидов. В 1966г. М.Ниренберг, Г. Корана и С. Очоа установили структуру всех 64 триплетов. 10. Структура гена прокариот Бактерии и вирусы по строению наследственного материала и способам его передачи занимают особое место в царстве природы. К ним в малой степени применимы закономерности наследования признаков, установленных для высших организмов при половом размножении. Они меньше защищены от воздействия внешней среды, что влечет за собой более высокую их изменчивость. В отличие от многоклеточных организмов у бактерий нет четко оформленного ядра. Его заменяют нуклеоид, который не отделен от остальной части клетки мембраной, как у эукариот. Вирусы подобно бактериям не одинаковы по форме. Однако для всех вирусов характерна более простая организация, чем для бактерий. Вне живой клетки вирусы не размножаются. Вирусы паразитирующие в бактериях называют – бактериофагами. Однако и между клетками разных штаммов бактерий и вирусов происходит обмен генетическим материалом, осуществляемый путем трансформации, трансдукции и конъюгации. Трансформация – это поглощение ДНК бактерии донора, в клетке ДНК бактерии реципиента. В процессе трансформации бактерий принимают участие 2 бактериальные клетки: донор и реципиент. Трансформирующий агент представляет собой часть молекулы ДНК донора, которая внедряется в геном реципиента. В процессе трансформации, клетки донора и реципиента не соприкасаются друг с другом. Из клеток донора выделяются в окружающую среду фрагменты молекул ДНК, которые адсорбируются на оболочке клетки реципиента, а в последствии втягиваются внутрь ее. Трансдукция – перенос гена из одной бактериальной клетки в другую при помощи бактериофага. Характерная особенность трансдукции заключается в том, что трансдуцированные бактерии приобретают только те свойства, которые были у донора. Для осуществления трансдукции необходимо присутствие бактерии – донора, бактерии реципиента и умеренного фага. Явление трансдукции установлено у многих бактерий. Как правило, трансдуцируется один ген, реже два и очень редко три сцепленных гена. Конъюгация – это перенос генетического материала от одной бактериальной клетки (донора) к другой (реципиенту) при их непосредственном контакте. При конъюгации бактерии сближались, клеточная оболочка в точке их соприкосновения растворялась, и между ними образовался цитоплазматический мостик, по которому хромосома одного штамма переходила в другой. Переход при конъюгации хромосомы от одной бактерии к другой происходит всегда от клеток одного определенного штамма к другому. При конъюгации целая хромосома донора переходит в клетку реципиента очень редко, чаще всего перемещается лишь участок ее. Таким образом, процесс конъюгации дает возможность изучить расположение генов в хромосоме бактерии. Структура гена прокариот сравнительно проста. Участок, кодирующий определенный белок, представляет ряд нуклеотидов (триплетных кодонов), которые транскрибируются на мРНК и затем транслируются на рибосоме в данный белок. Более сложной янляется система регуляции синтеза белка у бактерий. Как показали исследования Ф.Жакоба и Ж.Моно (1961), проведенные на E.coli, структурные гены, детерминирующие утилизацию этой бактерией лактозы, довольно тесно сцеплены и образуют оперон Это так называемая кластерная организация генов. При этом (см. рис. 12.1) происходит транскрипция и трансляция единой мРНК для генов Z, Y и А. Транскрипция инициируется геном-промотором (Р). Важная роль отведена гену-индуктору (lacl). Заметим, что ген-индук- тор может располагаться в другом месте генома, на удалении от оперо- на, активность которого он регулирует. Он кодирует белок /яс-репрсс- сор, который, связываясь в определенный момент с ДНК оператора (0+), репрессирует транскрипцию мРНК со структурных генов lacZ, lacY и lacA. Если белок-репрессор связывается с ДНК индуктора, репрессор становится неактивным и белок-репрессор отделяется от ДНК оператора, и тогда РНК-полимераза инициирует транскрипцию мРНК со структурных генов. Белок /яс-репрессор — аллостерический белок, который имеет два центра связывания и поэтому может присоединяться к двум разным молекулам. В одном случае он связывается с оператором, а в другом — с лактозой. 11. Особенности организации генома эукариот. Транскриптон У эукариот объем наследственного материала значительно больше. В отличие от прокариот в эукариотических клетках одновременно активно транскрибируется от 1 до 10% ДНК. Состав транскрибируемых последовательностей и их количество зависят от типа клетки и стадии онтогенеза. Значительная часть нуклеотидных последовательностей у эукариот не транскрибируется вообще — молчащая ДНК. Большой объем наследственного материала эукариот объясняется существованием в нем помимо уникальных также умеренно и высоко повторяющихся последовательностей. Высоко повторяющиеся последовательности ДНК располагаются в основном в гетерохроматине, окружающем центромерные участки. Они не транскрибируются. Умеренные повторы распределены по всему геному и транскрибируются в РНК. К ним относятся гены, контролирующие синтез гистонов, тРНК, рРНК и некоторые другие. Остальная часть генома представлена уникальными нуклеотидными последовательностями. Избыточность генома эукариот объясняется также экзон-интронной организацией большинства эукариотических генов, при которой значительная транскрибированной РНК удаляется в ходе следующего за синтезом процессинга и не используется для кодирования аминокислотных последовательностей. Избыточность генома эукариот объясняется также экзон-интронной организацией большинства эукариотических генов, при которой значительнаячасть транскрибированной РНК удаляется в ходе следующего за синтезом процессинга и неиспользуется для кодирования аминокислотных последовательностей. Транскриптон- структурно-функциональная единица эукариот. Регуляторные компоненты транскриптона: 1 Промотор - так же как и у оперона явялется местом начала транскрипции. Это участом молекулы ДНК с которым связывается РНК-полимераза 2 Модуляторы - участки транскриптона отвечающие за активность работы определенных генов, модуляторы подразделяются на: - сайленсеры - снижающие работу генов. - энхайсеры - ускоряющие работу генов. 3 Структурные гены которые представлены и интронами (некодирующими последовательностями) и экзонами (кодирующими последовательностями). 4 Терминатор - конечный участок оперона. Место завершения транскрипции. Логическая последовательность компонентов транскриптона: энхайсер, сайленсер, промотор, структурный ген. Особенность работы транскриптона состоит в том, что при «включении» сначала синтезируется про-иРНК, содержащая последовательности комплементарные как экзонам, так и интронам. Поэтому далее в ядре клетки происходит процессинг и-РНК– ферментативное разрушение неинформативной части и-РНК и расщепление информативной части на последовательности, комплиментарные экзонам. Далее из фрагментов информативной части и-РНК формируется моноцистронная и-РНК посредством сплайсинга (который может быть и альтернативным, что приводит к полиморфизму белков в конечном итоге) – сшивания фрагментов лигазами. Далее и-РНК выходит из ядра в рибосомы, происходит трансляция. Всё остальное происходитьпринципиально так же, как и в случае с опероном. 12. Классификация генов По функциям гены классифицируют на структурные и функциональные. Структурные гены содержат информацию о белках-ферментах, гистонах, о последовательности нуклеотидов в разных видах РНК. Функциональные гены оказывают влияние на работу структурных генов. Функциональными являются гены-модуляторы и гены-регуляторы. Гены-модуляторы – это ингибиторы, интенсификаторы, модификаторы. Они усиливают, ослабляют или изменяют работу структурных генов. Регулируют работу структурных генов гены-регуляторы и гены-операторы. Генотип всех соматических клеток организма одного вида одинаков. Но клетки разных тканей отличаются друг от друга. Вероятно, это связано с тем, что в них работают разные блоки генов. Область проявления действия данного гена называется полем его действия (например, распределение волосяного покрова на теле человека). Как правило, гены, детерминирующие определенные признаки, «работают» непостоянно (например, гены, определяющие синтез половых гормонов); их функция значительно снижается с возрастом. Период функционирования гена называется временем его действия. По месту действия гены подразделяют на три группы 1) функционирующие во всех клетках (например, гены, кодирующие ферменты энергетического обмена); 2) функционирующие в клетках одной ткани (детерминирующие синтез белка миозина в мышечной ткани); 3) специфичные для одного типа клеток (гены гемоглобина в незрелых эритроцитах). 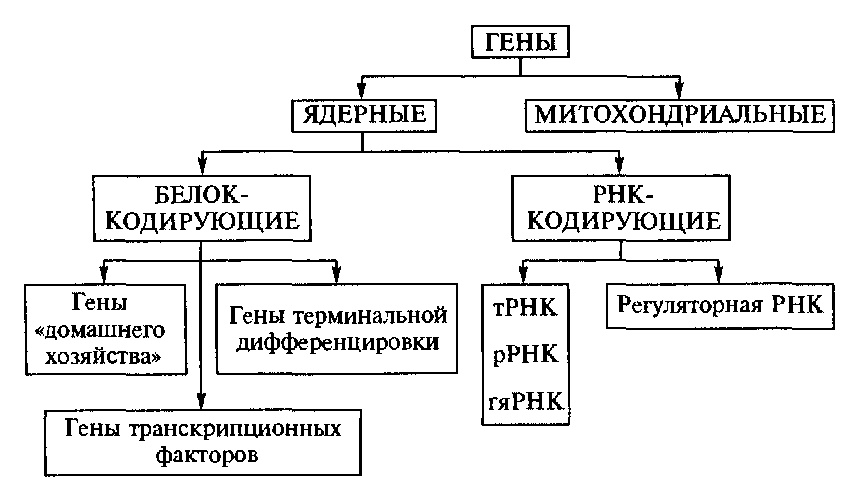 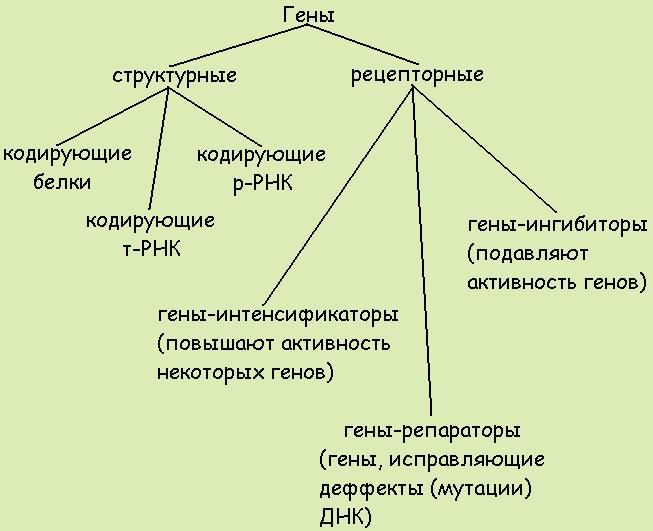 13. Основные классы нуклеотидных последовательностей в ДНК клеток человека, их биологическая роль 14. Уровни упаковки наследственного материала при подготовке клеток к делению Первый уровеньупаковки ДНК – нуклеосомный. Нуклеосомная нить хроматина (см. выше) имеет диаметр около 13 нм. После упаковки длина молекулы ДНК уменьшается в 5-7 раз. Нуклеосомный уровень обнаруживается в электронном микроскопе в интерфазе и при митозе. Второй уровень упаковки – соленоидный (супернуклеосомный). Нуклеосомная нить конденсируется, её нуклеосомы «сшиваются» гистоном Н1 и образуется спираль диаметром около 25 нм. Один виток спирали содержит 6-10 нуклеосом. Нить укорачивается еще в 6 раз. Супернуклеосомный уровень упаковки обнаруживается в электронном микроскопе как в интерфазных, так и в митотических хромосомах. 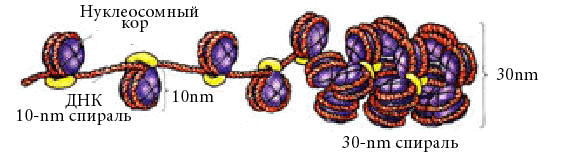 Третий уровень упаковки – хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель и изгибов. Она составляет основу хроматиды и обеспечивает хроматидный уровень упаковки. Он обнаруживается в профазе. Диаметр петель около 50 нм. Нить ДНП укорачивается в 10-20 раз. Третий уровень упаковки – хроматидный (петлевой). Супернуклеосомная нить спирализуется с образованием петель и изгибов. Она составляет основу хроматиды и обеспечивает хроматидный уровень упаковки. Он обнаруживается в профазе. Диаметр петель около 50 нм. Нить ДНП укорачивается в 10-20 раз. Четвертый уровень упаковки – уровень метафазной хромосомы. Хроматиды в метафазе способны спирализоваться с образованием эухроматиновых (слабо спирализованных) и гетерохроматиновых (сильно спирализованных) участков; происходит укорочение в 20 раз. Метафазные хромосомы имеют длину от 2,3 до 11 мкм и диаметр от 0,2 до 5,0 мкм. Общий итог конденсации – укорочение нити ДНП в 10000 раз. 15. Расположение генов в хромосомах Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу. Одним из классических опытов Моргана на дрозофиле, доказывающим линейное расположение генов, был следующий. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтую окраску тела y, белый цвет глаз w и вильчатые крылья bi, были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1,2% мух кроссоверных, возникших от перекреста между генами у и w; 3,5% − от кроссинговера между генами w и bi и 4,7% — между у и bi. Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между у и w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно. Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус. Основным положениям хромосомной теории наследственности — парности аллелей, их редукции в мейозе и линейному расположению генов в хромосоме — соответствует однонитчатая модель хромосомы. 16. Молекулярная организация хромосом Каждая хромосома состоит из одной или двух молекул ДНК (в зависимости от периода клеточного цикла), которые, связываясь с гистоновыми и негистоновыми белками, образуют нуклепротеиновые комплексы с разным уровнем конденсации: - нуклеосомный уровень- молекула ДНК с диаметром 2 нм взаимодействует с гистонами, формируя первичную нуклеосомную нить, единицей организации которой является гистоновый октамер из восьми молекул гистоновых белков (2Н2А, 2Н2В, 2НЗ и 2Н4) и ДНК; на каждый октамер приходится около двух витков спирали ДНК из «160 п.н.: в результате образуется нуклеопротеиновая полинуклеосоная нить диаметром 11 нм, а длина молекулы ДНК в результате упаковки сокращается в 6 раз; - соленоид – второй уровень упаковкигенетического материала; образуется посредством конденсации нуклеосомной нити при участий гистона HI; Диаметр нуклеосомной нити на этом уровне составляет около 30 нм, а уровень укладки достигает 40 раз; - третий уровень конденсации – петли, которые образуются следующим образом: соленоидная нить фиксируется к специальной продольной оси., образованной белками ядерного матрикса (scaffold); соленоид взаимодействует с осью в специфических сайтах (SAR) посредством специальных белков хромосомной нити (SAP); на этом уровне генетический материала конденсируется в 600 – 1000 раз; - четвертый уровень конденсации – метафазная хромосома- является результатом спирализации петель вокруг хромосомной оси после утери контакта scaffold / ядерная оболочка; один виток спирали состоит из примерно 30-ти петель, при этом обеспечивается максимальный (10 000 раз) уровень упаковки генетического материала. 17 Морфологические особенности метафазной хромосомы. На стадии метафазы митоза хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. У метафазных хромосом сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначают буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом. В зависимости от расположения центромеры различают три типа строения хромосом: - акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах; - субметацентрические хромосомы с плечами неравной длины; - метацентрические хромосомы, у которых центромера расположена посередине или почти посередине. Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка. Вторичные перетяжки бывают различной длины и могут располагаться в различных точках по длине хромосомы. Вторичные перетяжки отделяют от основной хромосомы так называемые спутники, содержащие многократные повторы генов, кодирующих рибосомные РНК. 18. Сателлитные и теломерные участки хромосом Теломе́ры — концевые участки хромосом. Теломерные участки хромосом характеризуются отсутствием способности к соединению с другими хромосомами или их фрагментами и выполняют защитную функцию. САТЕЛЛИТНАЯ ДНК (от лат. satelles — спутник, сообщник) — особая фракция ДНК в составе генома эукариот, которая отличается от основной ДНК по плотности. Сателлитная ДНК состоит из многократных линейных повторов основной короткой последовательности нуклеотидов и не транскрибируется. У современного человека обнаружены три фракции сателлитной ДНК. С их формированием предположительно связывают процесс эволюции мозга. |