3.2.5. Стебель — осевой орган побега
Общая характеристика стебля
Стеблем называют ось побега, несущую на себе листья, почки, цветы и плоды.
Основные функции:
опорная;
проводящая;
запасающая.
Дополнительные функции:
орган вегетативного размножения;
орган фотосинтеза.
Стебель, как и корень, развивается из зародыша семени, характеризуется радиальной симметрией и неограниченным ростом. Но в отличие от корня стебель обладает свойством отрицательного геотропизма.
Продолжительность жизни стеблей различна. У основной массы травянистых растений стебли живут в течение одного вегетационного периода (лен, гречиха, горох), у эфемеров и эфемероидов — от 2-6 недель до 5-6 месяцев (мокрица, бурачок пустынный, пролеска сибирская, тюльпаны). Наибольшая долговечность характерна для стеблей древесных растений: яблоня — до 200, сосна — до 500, дуб, ель — до 1200, кипарис — до 3000, секвойя — до 5000, драцена, баобаб — до 6000 лет.
Размеры стеблей колеблются в широких пределах: у ряски — несколько мм, а у ротанговой пальмы-лианы — до 300 м.
Выделяют два основных типа стебля:
деревянистый — обычно многолетний, утолщающийся неопределенно долго, образованный одревесневающими тканями (береза, виноград, смородина);
травянистый — обычно существующий один вегетационный период, как правило, не утолщающийся (или слабо утолщающийся) и неодревесневающий (или слабо одревесневающий) (крапива, лебеда).
Анатомия стебля
Анатомическое строение стебля более разнообразно, чем у корня. Оно может быть первичным и вторичным. Рассмотрим строение стебля древесного двудольного растения.
Стебли двудольных растений характеризуются следующими особенностями:
первичное строение рано сменяется вторичным;
кроме эпидермы, развиваются вторичные покровные ткани — перидерма и корка;
первичная кора хорошо развита и дифференцирована;
проводящие пучки расположены упорядоченно;
проводящие пучки открытого типа (имеют камбий).
Эпидерма, перидерма
и первичная кора
Рассмотрим строение древесного стебля двудольного растения на примере стебля липы (рис.21).
Молодые (однолетние) побеги липы покрыты эпидермой. На поперечном срезе заметны первичная кора, флоэма, камбий, вторичная ксилема, примыкающая к пер-
в

Рис. 21. Внутреннее строение стебля липы:
1 — эпидерма; 2 — пробка; 3 — первичная кора; 4 — луб; 5 — сердцевинный луч; 6 — камбий; 7 — осенний сосуд; 8 — весенний сосуд; 9 — первичная древесина; 10 — сердцевина; 11 — чечевичка.
ичной ксилеме, окружающей сердцевину. Уже летом под эпидермой закладывается пробковый камбий — феллоген. Он откладывает наружу клетки пробки, а внутрь — клетки феллодермы. Пробка, феллоген и феллодерма образуют общий вторичный покров — перидерму. Под некоторыми устьицами закладываются чечевички. У двух-трехлетней ветви липы под перидермой находятся кора (первичная и вторичная), камбий, древесина и сердцевина.
Камбий
Большая часть стебля образована тканями,
возникшими в результате деятельности камбия, располагающегося между вторичной корой и древесиной. Именно благодаря ему происходят вторичные изменения в строении стебля. (соотношение 4:1 соответственно Обычно внутрь камбий откладывает большее число производных, чем наружу). Весной клетки камбия активно делятся, с приближением осени деятельность камбия ослабевает, и зимой он вступает в период покоя.
Кора
Все ткани, лежащие кнаружи от камбия, называют
корой, которая образована флоэмой (лубом) и сердцевинными лучами. На срезах сердцевинные лучи имеют вид светлых треугольников. Они чередуются с трапецевидными участками флоэмы. В виде узких лент сердцевинные лучи продолжаются в ксилеме (древесине). По ним в радиальном направлении осуществляется передвижение питательных веществ. К осени в них накапливаются крахмал и масло, которые расходуются весной.
Состав флоэмы липы неоднороден. Луб образован ситовидными трубками, клетками-спутницами, лубяной паренхимой и лубяными волокнами. По ситовидным трубкам происходит передвижение веществ, образованных в листьях в процессе фотосинтеза. В лубяной паренхиме накапливаются питательные вещества, которые весной гидролизуются и расходуются растением. Лубяные волокна обеспечивают прочность.
Древесина
Внутрь от камбия откладываются клетки древесины (вторичной ксилемы), в состав которой входят сосуды, трахеиды, древесная паренхима и древесная склеренхима (волокна). Особенностью ксилемы является одревеснение клеточных стенок (за исключением клеток древесной паренхимы). Вторичная ксилема составляет основную массу (9/10 объема) древесного стебля.
В результате периодической деятельности камбия в древесине образуются
годичные кольца — прирост древесины за один вегетационный период. Весной камбий откладывает широкопросветные и тонкостенные сосуды и трахеиды, к которым примыкают клетки древесной паренхимы. Осенью в древесине преобладают узкопросветные и толстостенные сосуды, трахеиды и древесные волокна. Переход от весенней древесины к осенней постепенный, от осенней к весенней всегда резкий. Весенняя древесина более светлая, чем осенняя. Поэтому между годичными кольцами возникает хорошо заметная граница. По годичным кольцам можно определить возраст дерева. Ширина годичных колец неодинакова: в благоприятные годы образуются более широкие кольца, чем в неблагоприятные. У тропических растений, растущих непрерывно в течение года, годичные кольца не образуются.
Сердцевина
В центре стебля находится
сердцевина, образованная округлыми паренхимными клетками. Она окружена небольшим количеством сосудов первичной ксилемы.
Транспорт веществ по стеблю
Одна из основных функций стебля — транспорт воды, минеральных и органических веществ.
Транспорт воды и минеральных веществ
Вода и минеральные вещества поглощаются растением из почвы корнями поступают в сосуды и трахеиды ксилемы стебля. (Механизм транспорта см. в теме "Корень").
Транспорт органических веществ
Главное направление перемещения метаболитов — отток продуктов фотосинтеза (
ассимилятов) от листьев в другие органы растения. Функцию транспорта органических веществ выполняет флоэма, основные элементы которой (ситовидные трубки и клетки-спутницы) образуют единую транспортную систему.
Доказательством транспорта органических веществ служат опыты с "кольцеванием" стеблей древесных растений, впервые проведенные в XVII итальянцем Марчелло Мальпиги. Если на фотосинтезирующем дереве срезать кольцо коры, то над ним будет происходить образование утолщения (кора набухает). Это свидетельствует о накоплении продуктов ассимиляции, передвигающихся от листьев вниз по флоэме.
По флоэме растворенные органические вещества перемещаются в двух направлениях — вверх и вниз (в отличие от ксилемы, по которой вода и минеральные вещества транспортируются только вверх). По-видимому, одновременный разнонаправленный ток органических веществ осуществляется по разным ситовидным трубкам.
3.2.6. Лист — боковой орган побега
Лист — боковой (латеральный) орган, характеризующийся ограниченным ростом.
Как правило, лист — плоский дорсовентральный (разносторонний) орган: верхняя (брюшная) и нижняя (спинная) стороны построены различно.
Лист растет только в ограниченный период времени своим основанием путем вставочного роста (однодольные) или всей поверхностью (двудольные).
У большинства растений лист живет, как правило, в течение одного вегетационного периода, а у вечнозеленых — 1-5 лет (иногда 10-15 лет, как у ели, араукарии). У однолетних растений продолжительность жизни листа примерно равна длительности жизни стебля, у многолетних — значительно короче.
Главные функции листа:
фотосинтез;
газообмен;
транспирация.
Дополнительные функции листа:
запасающая (сочные чешуи луковицы);
вегетативное размножение (бегония);
защитная (колючки кактуса).
М
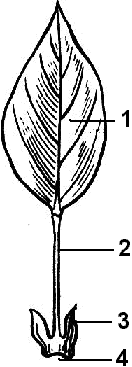
Рис. 22. Внешнее строение листа:
1 — листовая пластинка; 2 — черешок; 3 — прилистники; 4 — основание листа.
орфология листа
Основные части листа
Лист большинства растений состоит из пластинки, черешка, прилистников основания (рис.22).
Листовая пластинка
Листовая пластинка — расширенная, обычно плоская часть листа, выполняющая функции фотосинтеза, транспирации и газообмена.
По форме листовые пластики бывают: округлые (настурция), овальные (лещина древовидная), элиптические (яблоня лесная), яйцевидные (сирень), линейные (пшеница), ланцетные (ива) и т.д. В основе классификации листьев по форме листовой пластинки лежит соотношение длины и ширины пластинки и местоположение наибольшей ширины. Отдельно выделяют также:
стреловидный лист — лист, у которого боковые лопасти в нижней части листовой пластинки расходятся под острым углом к черешку (стрелолист);
копьевидный лист — лист, у которого лопасти отходят под прямым или тупым углом (щавель).
Ф
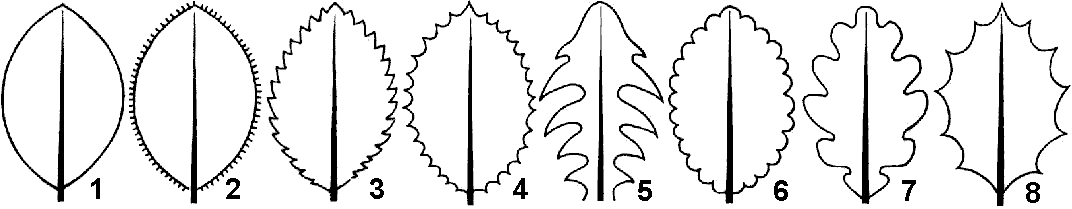
Рис. 23. Форма края листовой пластинки:
1 — цельнокрайный; 2. — реснитчатый; 3 — пильчатый; 4 зубчатый; 5 — струговидный; 6 — городчатый; 7 — волнистый; 8 — выемчатый.
орма края листовой пластинки (рис. 23) бывает: цельнокрайной (сирень), зубчатой (крапива), пильчатой (липа), городчатой (будра), выемчатой (осина) и т.д.
Если надрезанность края не превышает одной четверти ширины полупластинки, то листья называют
цельными (сирень, береза). Если же надрезанность пластинки больше, то такие листья называются
расчлененными (смородина, одуванчик). По степени расчленения листовой пластинки различают (рис. 24):
л

Рис. 24. Расчленение листовой пластинки:
1 — перисто-лопастной; 2 — перисто-раздельный; 3 — перисто-рассеченный; 4 — пальчато-лопастной; 5 — пальчато-раздельный; 6 — пальчато-рассеченный.
опастные листья — выемки не доходят до половины полупластинки (огурец, дуб);
раздельные листья — выемки заходят глубже половины полупластинки (лютик, герань);
рассеченные листья— выемки достигают главной жилки листа (картофель, гусиная лапка).
Расчленение листовой пластинки может быть пальчато-лопастные (клен), пальчато-раздельные (клещевина) и пальчато-рассеченные (лютик) листья, а также перисто-лопастные (дуб), перисто-раздельные (полынь) и перисто-рассеченные (тысячелистник).
Черешок
Черешок — суженная часть листа, соединяющая листовую пластинку с основанием и регулирующая положение листа по отношению к свету. Листья с черешками называют
черешковыми, без черешков —
сидячими.
Основание листа
О
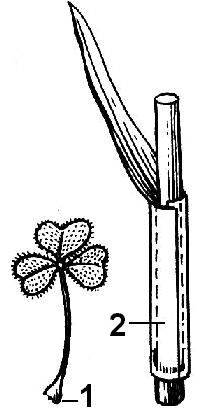
Рис. 25. Основание листа:
1 — листовая подушечки; 2 — влагалище
снование листа — нижняя часть листа, примыкающая к стеблю (рис. 25). Часто слабо развита. Защищает верхушечные и пазушные почки во время их развития. Одной из форм является
листовое влагалище — расширенное основание листа в виде трубки, охватывающее часть стебля (пшеница, лук-порей).
Прилистники
Прилистники — листовидные образования у основания листа, которые служат для защиты молодого листа и пазушной почки. Встречаются не у всех растений. У большинства растений у взрослых листьев прилистники отсутствуют (дуб, береза, черемуха). Иногда прилистники достигают значительного развития, их размеры превышают размеры листовых пластинок (горох). В этом случае прилистники выполняют роль фотосинтезирующих органов.
Листорасположение
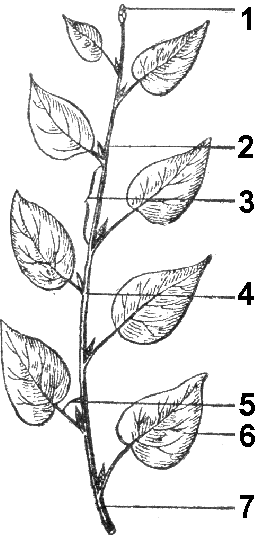
Листорасположение — порядок размещения листьев на оси побега (рис. 26). Может быть:
о
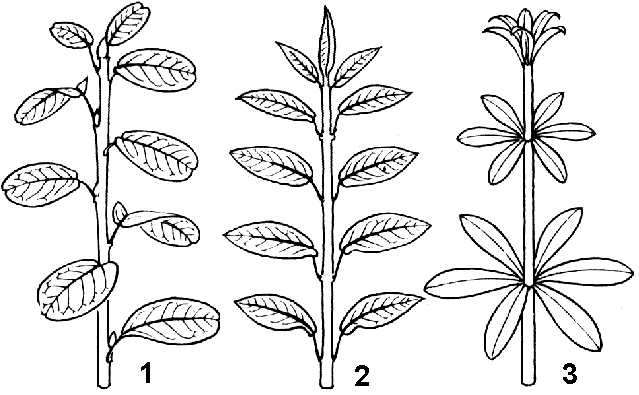
Рис. 26. Листорасположение:
1 — очередное; 2 — супротивное; 3 — мутовчатое.
чередное, или спиральное — на каждом узле располагается по одному листу, причем основания листьев можно соединить условной спиральной линией, растянутой вдоль оси побега (береза, липа);
супротивное — от узла отходят два сидящих друг против друга листа (клен, сирень);
мутовчатое — от узла отходит более двух листьев (олеандр, вороний глаз).
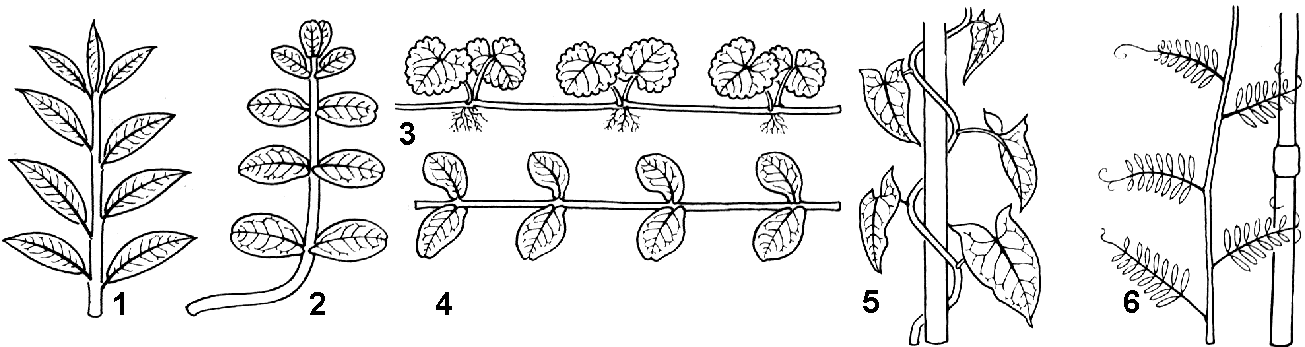
Классификация листьев
Различают листья простые и сложные. Листья, имеющие одну пластинку (цельную или выемчатую), называются
простыми. Простые листья при
л
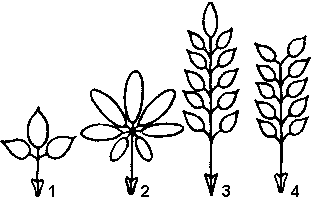
Рис. 27. Сложные листья:
1 — тройчатосложный; 2 — пальчатосложный; 3 — непарноперистосложный; 4 — парноперистосложный.
истопаде опадают целиком или вообще не опадают (у большинства травянистых растений). Такие листья свойственны подавляющему большинству растений (береза, клен, одуванчик).
Сложные листья- — листья, состоящие из нескольких четко обособленных листовых пластинок (листочков), каждый из которых своим черешком прикреплен к общему черешку (рахису). Часто сложный лист опадает по частям: сначала листочки, а потом черешок.
В зависимости от расположения листочков различают (рис. 27):
Перистосложные листья — листья, у которых листочки располагаются по бокам рахиса. Когда верхушка рахиса заканчивается одним непарным листочком, такие листья называются непарноперистыми (шиповник, акация белая). У парноперистого листа все листочки имеют себе пару (горох, акация желтая).
Пальчатосложные листья — листья, у которых листочки располагаются не по длине рахиса, а лишь на его верхушке в одной плоскости (каштан, люпин).
Частным случаем сложного листа является
тройчатый лист — лист, имеющий только три листочка (клевер, кислица).
Рахис сложных листьев может образовывать боковые ответвления, тогда возникают дважды- , трижды- четыреждыперистосложные листья. Например, у мимозы дваждыперистосложный лист.
Жилкование листьев
Жилкование — это система проводящих пучков в листовых пластинках.
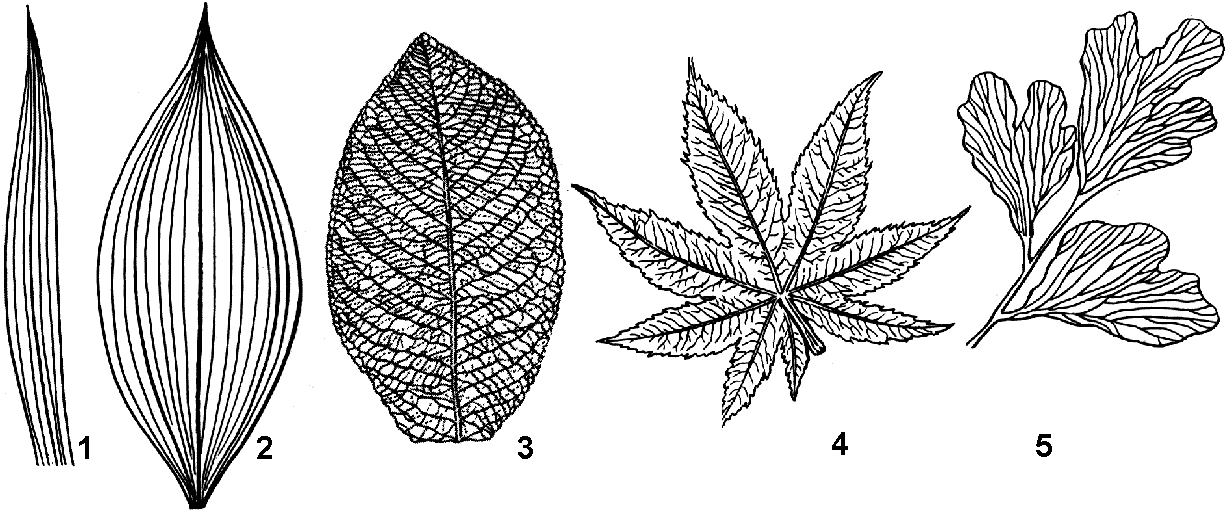
Рис. 28. Жилкование листьев:
1 — параллельное; 2 — дуговое; 3 — сетчатое с перистым расположением основных жилок; 4 — сетчатое с пальчатым расположением основных жилок; 5 — дихотомическое.
Характер расположения жилок и форма листовых пластинок тесно взаимосвязаны (рис. 28). Различают:
простое жилкование — листовую пластинку от основания до верхушки пронизывает только одна жилка (мхи, плауны);
дихотомическое жилкование — листовую пластинку пронизывают вильчато разветвленные жилки (гинкго);
дуговое жилкование — листовую пластинку от основания до верхушки пронизывает несколько одинаковых жилок, располагающихся дугообразно (ландыш, чемерица);
параллельное жилкование — листовую пластинку от основания до верхушки пронизывает несколько одинаковых жилок, располагающихся строго параллельно (рожь, осока);
сетчатое жилкование — обычно из черешка в листовую пластинку входит одна жилка, которая затем дает ответвления — боковые жилки, образующие густую сеть. Сетчатое жилкование может быть перистым и пальчатым.
Анатомия листа
Микроскопическое строение листа сходно у многих растений (рис. 29). На поперечном срезе листовой пластинки видно, что сверху и снизу лист покрыт эпидермой (кожицей). Поверх эпидермы располагается слой кутина. Как правило, на верхней эпидерме почти полностью отсутствуют устьица. Нижняя поверхность листа покрыта эпидермой с множеством устьиц. На 1 мм
2 листа приходится от 50 до 500 устьиц. У плавающих на поверхности воды листьев устьица располагаются на верхней эпидерме, а у погруженных листьях обычно отсутствуют.
М

Рис. 29. Внутреннее строение листа:
1 — кутикула; 2 — эпидерма; 3 — ксилема; 4 — флоэма; 5 — волокна; 6 — колленхима; 7 — устьица; 8 — столбчатая хлоренхима; 9 — губчатая хлоренхима; 10 — железистый волосок; 11 — кроющий волосок; 12 — межклетник.
ежду верхней и нижней эпидермой располагается мезофилл, образованный столбчатой и губчатой хлоренхимой. Столбчатая хлоренхима располагается под верхней кожицей листа. В основном в ней осуществляются процессы фотосинтеза. У растений средних широт (огурец, клевер) столбчатая паренхима обычно образована одним рядом клеток, у южных растений чаще образуется 2-3 ряда.
Ближе к нижней эпидерме располагается губчатая хлоренхима,
осуществляющая преимущественно функции газообмена и транспирации. Клетки губчатой хлоренхимы принимают участие и в фотосинтезе, но в меньшей слабой степени, чем клетки столбчатой паренхимы, так как число хлоропластов в них в 2 -6 раз меньше.
Жилки образуют проводящую систему листа. В жилках имеются проводящие пучки (одним или несколькими). Большинство пучков закрытые, лишь более крупные могут быть отрытыми, но камбий развит слабо. Камбиальная активность наиболее сильно выражена у вечнозеленых растений.
В проводящих пучках ксилема обращена к верхней стороне листа, а флоэма — к нижней. Крупные проводящие пучки образованы сосудами и ситовидными трубками. В мелких пучках сосуды сменяются трахеидами, а ситовидные трубки — паренхимными клетками. Обычно проводящие пучки окружены обкладкой из паренхимы —
обкладочной паренхимой. Обкладка увеличивает площадь контакта мезофилла с проводящими элементами ксилемы и флоэмы.
Проводящие пучки выполняют и механическую функцию. Крупные жилки, как правило, окружены склеренхимой.
Функции листа
Фотосинтез
Одна из основных функций листа — фотосинтез. Сущность фотосинтеза заключается в том, что зеленые растения поглощают солнечную энергию и из воды и углекислого газа при участии минеральных веществ создают сложные органические соединения.
Фотосинтез — сложный, многоступенчатый процесс. Он представляет собой последовательную цепь окислительно-восстановительных реакций, часть которых идет с непосредственным использованием света, другая же света не требует. Таким образом, весь процесс фотосинтеза можно разделить на две фазы: световую и темновую.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит активирование молекул хлорофилла за счет энергии солнечного света. Избыточная энергия молекул хлорофилла используется на синтез АТФ и фотолиз (расщепление) воды на ионы Н
+ и ОН
-. Эти процессы носят ферментативный характер и происходят с участием системы переносчиков электронов. Атомы водорода участвуют в восстановлении НАДФ, а ионы ОН
-, взаимодействуя друг с другом, образуют молекулярный кислород и воду. Молекулярный кислород выделяется в атмосферу, а АТФ и НАДФ
.Н
2 используются для восстановления углерода в темновой фазе фотосинтеза.
Темновая фаза фотосинтеза
Темновая фаза фотосинтеза включает реакции, идущие за счет энергии, накопленной в световую фазу или при дыхании. В темновой фазе фотосинтеза выделяют несколько этапов:
фиксация СО2 рибулозобифосфатом;
перенос активного водорода на соединение, в котором фиксирована поглощенная молекула СО2;
восстановление акцептора СО2;
образование сахаров.
(Подробнее о процессе фотосинтеза см. раздел "Общая биология").
Дыхание
Дыхание — одна из важнейших сторон обмена веществ. Оно служит источником энергии, необходимой для осуществления всех процессов жизнедеятельности растения.
К

Рис. 30. Дыхание листа.
ак и любой орган, лист интенсивно дышит, то есть поглощает кислород и выделяет углекислый газ. Причем процесс дыхания осуществляется постоянно, как на свету, так и в темноте. Если поместить в сосуд свежесрезанные листья, плотно закрыть его и поставить в темное теплое место, то на следующий день можно обнаружить, что состав воздуха в сосуде изменился. Если поместить в него свечу, то она погаснет, а известковая вода мутнеет.
Этот простой опыт доказывает, что в сосуде
уменьшилось количество кислорода и увеличилось количество углекислого газа, то есть листья дышат (рис. 30).
В отличие от фотосинтеза, во время дыхания происходит освобождение энергии, органические вещества окисляются, и происходит выделение углекислого газа.
Дыхание представляет собой цепь окислительно-восстановительных реакций, катализируемых ферментами. Эти реакции протекают в митохондриях клеток. (Механизм клеточного дыхания см. раздел "Общая биология").
Транспирация
Под
транспирацией понимают испарение воды листьями (рис. 31). Транспирация обеспечивается ря-
д
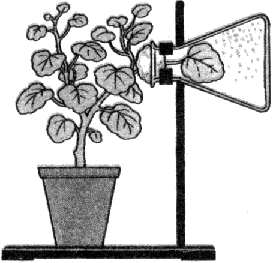
Рис. 31. Испарение воды листьями.
ом анатомических и физиологических механизмов и поэтому является физиологическим процессом, в отличие от чисто физического испарения воды.
Значение транспирации очень велико, так как обеспечивает
терморегуляцию растения;
деятельность верхнего концевого двигателя водного тока.
Различают два вида транспирации — кутикулярную и устьичную.
Кутикулярная транспирация
Кутикулярная транспирация представляет собой процесс испарения воды с поверхности кутикулы, покрывающей лист. Кутикулярная транспирация составляет порядка 10-20% общего испарения воды взрослыми листьями.
Устьичная транспирация
Под
устьичной транспирацией понимают процесс испарения воды листьями с помощью особых структур — устьиц. Устьичная транспирация является основным механизмом водообмена между растением и атмосферой.
Процесс устьичной транспирации можно разделить на 3 основные этапа:
испарение воды с поверхности клеток в межклетники;
выход паров воды из межклетников через устьичную щель;
диффузия паров воды от поверхности листа в более далекие слои атмосферы.
В настоящее время общепризнана гипотеза устьичных движений, связанная с перераспределением ионов калия между замыкающими и сопутствующими клетками, приводящим к перемещению воды. Причем основную роль в создании осмотического давления в замыкающих клетках играют ионы калия, которые закачиваются в них, и образующаяся на свету глюкоза.
Определенную роль играет и концентрация СО
2. Избыток СО
2, по-видимому, вызывает подкисление цитоплазмы. Это приводит к изменению рН, что приводит к закрыванию устьиц.
Таким образом, устьица чутко реагируют на изменения внешних условий и физиологические изменения в тканях листа, что влияет на интенсивность транспирации, приспосабливая ее к конкретным условиям.
При открывании устьиц углекислота поступает в лист и создает предпосылки для фотосинтеза. В отсутствие света фотосинтез в замыкающих клетках прекращается (как и во всех других), тургорное давление снижается и устьица закрываются. При недостатке поступления воды в растение устьица тоже закрываются, сберегая таким образом то небольшое количество влаги, которое доступно растению.
Видоизменения листа
Лист — один из самых пластичных органов растения. В процессе приспособления к условиям среды обитания у всего листа или его части может произойти смена главной функции. Это приводит к качественным изменениям во внешнем облике и внутреннем строении листа, то есть возникают видоизменения или метаморфозы листа (рис. 32).
Колючки
Это видоизменение характерно для растений, обитающих в сухом и жарком климате, хотя нередко они возникают и у растений других климатических зон. Колючки уменьшают транспирацию и защищают растения от поедания животными.
Метаморфоз всего листа в колючку характерен для кактусов. У многих астрагалов, эспарцетов в колючку превращается рахис сложного листа, у белой акации — прилистники.
Усики
Это нитевидные образования, чувствительные к прикосновению и приспособленные для лазания. У вики, чечевицы, гороха в усик преобразуются верхняя часть рахиса и несколько верхних листочков. У чины безлистной все листочки редуцируются, а рахис представляет собой единственный усик (функцию фотосинтеза берут на себя прилистники.
Ловчие аппараты
Встречаются у растений, произрастающих на болотистых, торфяных, бедных минеральными веществами почвах. При помощи ловчих аппаратов росянка в средней полосе, венерина мухоловка в Северной Америке, непентес в тропической Азии используют богатую
азотом и фосфором органическую пищу, переваривая животных. Строение ловчих аппаратов различно (кувшинчики непентесов, ловушки венериной мухоловки, листья росянки), но все они способны улавливать и переваривать насекомых и других мелких животных с помощью ферментов, выделяемых особыми пищеварительными железками.
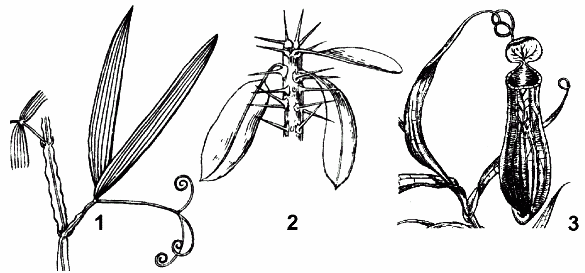
Рис. 32. Видоизменения листа:
1 — усик чины безлистной; 2 — колючка молочая блестящего; 3 — ловчий аппарат непентеса.
Листопад
При достижении листьями предельных размеров в них начинаются процессы старения, приводящие к гибели листа. По мере старения в листьях снижается интенсивность фотосинтеза и дыхания, уменьшается содержание в тканях белкового азота и РНК. Начинают преобладать процессы распада, а не синтеза. Органические вещества оттекают из старых тканей листа. Одновременно с этим в листьях накапливаются некоторые соли, например кристаллы оксалата кальция.
Верным признаком старения листа является изменение его окраски. Листья теряют зеленую окраску в результате деградации хлоропластов. Пожелтение и покраснение листьев связано с накоплением в них каротиноидов и антоциана.
У однодольных и травянистых двудольных листья постепенно отмирают и разрушаются, оставаясь на стеблях. У деревьев и кустарников
л

Рис. 33. Образование отделительного слоя:
1 — древесина; 2 — сердцевина; 3 — луб; 4 — перидерма; 5 — отделительный слой; 6 — проводящий пучок; 7 — черешок листа.
истья опадают. Массовое опадание листьев называют
листопадом. Опадание листьев обусловлено изменениями, происходящими в листе, а именно в месте прикрепления листа к стеблю. У основания черешка в поперечном направлении образуется специальный отделительный слой, состоящий из легко расслаивающейся паренхимы (рис. 33). Со стороны стебля ближайшие к основанию черешка клетки пробковеют и образуют защитный слой, сохраняющийся после опадания листа в виде листового рубца. Некоторое время лист держится за счет жилок. Но под действием силы тяжести листа и порывов ветра они разрываются, и листья опадают.
 Скачать 34.42 Mb.
Скачать 34.42 Mb.