Карташев Наумов Практикум по зоологии. "Практикум по зоологии позвоночных". Подтип бесчерепные acrania тема строение ланцетника систематическое положение объекта
 Скачать 5.56 Mb. Скачать 5.56 Mb.
|

|
5); у хвостатых земноводных к их концам прикрепляются коротенькие ребра, у бесхвостых земноводных ребра отсутствуют. Позвонки соединяются друг с другом сочленением самих тел позвонков (что обеспечивается процельным типом их строения) и соединением специальных парных сочленовных отростков (processus atricularis; рис. 55, 6), располагающихся спереди и сзади на основании верхней дуги. По сравнению с рыбами для земноводных характерна большая дифференцировка позвоночного столба на отделы, изменение формы тел позвонков и более сильное развитие сочленовных отростков. Эти преобразования связаны с наземным образом жизни и обеспечивают большую прочность осевого скелета при сохранении его подвижности, прочное соединение с ним тазового пояса и допускают некоторую подвижность черепа в вертикальной плоскости относительно туловища (возможность поднимать и опускать голову). Череп. Осевой, или мозговой, череп земноводных, как и череп хрящевых рыб, платибазального типа: с широким основанием и широко расставленными глазницами, между которыми располагается передний конец головного мозга. В черепе, по сравнению с костистыми рыбами, сохраняется много хряща, а число окостенений относительно невелико. 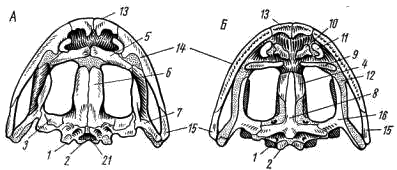 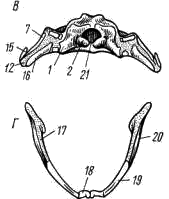 Рис. 56. Череп лягушки. А — сверху; Б — снизу; В — сзади; Г — нижняя челюсть сверху, пунктиром показаны хрящевые участки черепа 1 — боковая затылочная кость, 2 — затылочный мыщелок, 3 — переднеушная кость, 4 — клиновидно-обонятельная кость, 5 — носовая кость, 6 — лобно-теменная кость, 7 — чешуйчатая кость, 8 — парасфеноид, 9 — нёбная кость, 10 — сошник, 11 — хоана, 12 — нёбно-квадратный хрящ, 13 — межчелюстная кость, 14 — верхнечелюстная кость, 15 — квадратно-скуловая кость, 16 — крыловидная кость, 17 — меккелев хрящ, 18 — подбородочно-челюстная кость, 19 — зубная кость, 20 — угловая кость, 21 — большое затылочное отверстие В хряще затылочного отдела мозгового черепа образуются только парные боковые затылочные кости (occipitale laterale; рис. 56, 1), окаймляющие большое затылочное отверстие (foramen occipitale magnum; рис. 56, 21). Каждая из них образует мыщелок (condylus occipitalis; рис. 56, 2) для сочленения с шейным позвонком. В области слуховой капсулы вместо пяти пар ушных костей, характерных для костистых рыб, у земноводных возникает лишь одна пара — переднеушные кости (prooticum; рис. 56, 3). В передней части мозгового черепа при окостенении хряща образуется непарная клиновиднообонятельная кость (sphenethmoideum, рис. 56, 4), имеющая вид костного кольцевого пояска. 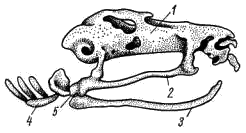 Рис. 57. Хрящевой череп головастика: 1 — мозговой череп, 2 — нёбно-квадратный хрящ, 3 — меккелев хрящ, 4 — жаберные дуги, 5 — челюстной сустав Вся остальная часть мозгового черепа остается хрящевой. Ее укрепляют покровные (кожные) кости. Сверху в передней части черепа лежат имеющие удлиненно- треугольную форму парные носовые кости (nasale; рис. 56, 5), затем слившиеся из лобных и теменных костей парные лобно-теменные кости (frontoparietale, рис. 56, 6), и кнаружи от ушных костей — имеющие сложную форму чешуйчатые кости (squamosum; рис. 56, 7). Дно мозгового черепа прикрывает мощная покровная кость крестообразной формы — парасфеноид (parasphenoideum; рис. 56, 8). Спереди от него лежат тоже покровные парные нёбные кости (palatinum; рис. 56, 9) и парные сошники (vomer; рис. 56, 10); на сошниках сидят мелкие зубы. Впереди сошников располагаются парные внутренние ноздри — хоаны (рис. 56, 11). Висцеральный отдел черепа земноводных также сохраняет много хряща. В течение всей жизни сохраняется нёбно-квадратный хрящ (cartilago palatoquadratum; рис. 56, 12), прирастающий передним концом к обонятельной области мозгового черепа, а задним — к основанию черепа перед слуховой капсулой (рис. 57, 2). Поэтому череп земноводных, как и остальных наземных позвоночных животных, по типу прикрепления челюстной дуги — аутостилический. К нёбно-квадратному хрящу прилегают возникающие в коже кости вторичной верхней челюсти: парные межчелюстные кости (intermaxillare или praemaxillare; рис. 56, 13), несущие зубы и верхнечелюстные кости (maxillare; рис. 56, 14). За ними, укрепляя заднюю часть нёбно-квадратного хряща, сверху образуется покровная квадратно- скуловая кость (quadratojugale; рис. 56, 15), а снизу также покровная — крыловидная кость (pterygoideum; рис. 56, 16). Первичная нижняя челюсть — меккелев хрящ (cartilago Meckeli; рис. 56, 17) также остается хрящевой, только самый ее передний конец окостеневает в маленькие парные подбородочно-челюстные кости (mento-mandibulare, рис.56, 18). К ним присоединяются покровные зубные кости (dentale; рис. 56, 19), у земноводных лишенные зубов. Задняя часть меккелева хряща обрастает длинной покровной угловой костью (angulare; рис. 56, 20) и еще несколькими мелкими покровными косточками. Через суставной отросток меккелева хряща нижняя челюсть подвижно сочленяется с задней частью нёбно- квадратного хряща (рис. 57, 5). 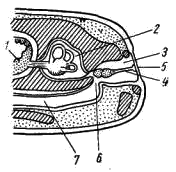 Рис. 58. Схематический разрез через слуховую область головы лягушки: 1 — головной мозг, 2 —слуховая капсула с полукружными каналами, 3 — полость среднего уха, 4 — стремечко, 5 — барабанная перепонка, 6 — евстахиева труба, 7 — ротовая полость Полная редукция у земноводных жаберной крышки и смена гиостилического типа прикрепления челюстей аутостилистическим приводят к потере основных функций подъязычной дуги (укрепление челюстей, опора жаберной крышки). Подъязычная дуга еще у предков современных земноводных начала редуцироваться, а полость брызгальца (остаток жаберной щели между челюстной и подъязычной дугами) в связи с переходом к жизни в воздушной среде преобразовалась в полость среднего уха (рис. 58, 3). Расположенный рядом с брызгальцем верхний элемент подъязычной дуги — подвесок (hyomandibulare) превратился в слуховую косточку—столбик, или стремечко (columel1а, или stapes; рис. 58, 4). У современных бесхвостых земноводных стремечко имеет вид тоненькой палочковидной косточки, лежащей перпендикулярно мозговому черепу под чешуйчатой и квадратно-скуловой костями. Одним концом стремечко упирается в центр барабанной перепонки (рис. 58, 5), а другим — в овальное окно слуховой капсулы. Этот механизм, усиливающий звуковые колебания и обеспечивающий возможность слуха в воздушной среде, у части современных земноводных в различной степени вторично редуцирован. Добавочными механизмами, обеспечивающими восприятие звуковых волн, распространяющихся по твердому субстрату, у них становится нижняя челюсть, а также передача звуковых колебаний по кровеносным стволам. 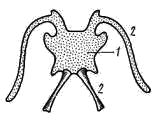 Рис. 59. Подъязычный аппарат лягушки: 1 — тело, 2 — рожки Нижний элемент подъязычной дуги — гиоид (hyoideum) и функционирующие у личинок земноводных жаберные дуги во время метаморфоза превращаются в подъязычный аппарат (рис. 59). У бесхвостых земноводных он представляет собой хрящевую пластинку с двумя главными нарами отростков — рожек. Передние, более длинные рожки (видоизменившиеся гиоиды) направляются назад и вверх и прикрепляются к стенкам слуховых капсул мозгового черепа. Подъязычный аппарат укрепляет дно ротовой полоски: к нему прикрепляются мышцы, расположенные между ветвями нижней челюсти. Предполагают, что гортанные хрящи также представляют собой преобразованные остатки жаберных дуг. Парные конечности и их пояса. Конечности земноводных, кяк и конечности других классов наземных позвоночных животных, представляют в схеме систему рычагов, подвижно соединенных друг с другом. Схемы строения передней и задней конечностей однотипны (рис. 60): 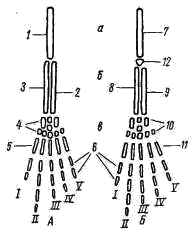 Рис. 60. Схема строения парных конечностей (левых) наземных позвоночных. А — передняя конечность; Б — задняя конечность: а — плечо — бедро, б — предплечье — голень, а — кисть—стопа; 1 — плечевая кость, 2 — локтевая кость, 3 — лучевая кость, 4 — запястье, 5 — пясть, 6 — фаланги пальцев, 7 — бедренная кость, 8 — большая берцовая кость, 9 — малая берцовая кость, 10 — предплюсна, 11 — плюсна, 12 — коленная чашечка, I — V — пальцы Проксимальный отдел передней конечности — плечо (humerus; рис. 61, 7) — трубчатая кость; средняя ее часть называется диафизом, а утолщенные концы — эпифизами. У земноводных эпифизы плеча (и бедра) остаются хрящевыми. Проксимальный конец имеет округлую головку плеча (caput humeri; рис. 61, 2), которая входит в суставную ямку пояса передних конечностей; на дистальном конце — полушаровидная поверхность для сочленения с костями предплечья. Поверхность плеча имеет гребни, к которым прикрепляются мышцы. У бесхвостых земноводных лежащая снаружи локтевая (ulna; рис. 61, 4) и с внутренней стороны — лучевая (radius; рис. 61, 5) кости сливаются в единую кость предплечья (antebrachium, рис. 61, 3); продольная бороздка показывает границу их слияния. У хвостатых земноводных эти кости самостоятельны. Проксимальные концы обеих костей образуют сочленовную ямку для соединения с плечом; позади этой ямки находится локтевой отросток (рис. 61, 6) локтевой кости, ограничивающий разгибание конечности. Запястье (carpus, рис. 61, 7) состоит из двух рядов мелких костачек. К дистальному ряду косточек запястья примыкают пять удлиненных косточек пясти (metacarpus; рис. 61, 8). С дистальными концами пястных костей сочленяются фаланги пальцев (phalanges digitorum; рис. 61, 9). У земноводных первый (большой) палец сильно редуцирован и кисть заканчивается лишь четырьмя хорошо развитыми пальцами. .
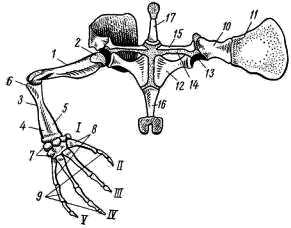 Рис. 61. Передняя конечность и плечевой пояс лягушки: 1 — плечевая кость, 2 — головка плеча, 3 — предплечье, 4 — локтевая кость, 5 — лучевая кость, 6 — локтевой отросток, 7 — запястье, 8 — пясть, 9 — фаланги пальцев, 10 — лопатка, 11 — надлопаточный хрящ, 12 — коракоид, 13 — суставная впадина для головки плеча, 14 — прокоракоидный хрящ, 15 — ключица, 16 — грудина, 17 — предгрудинник, I — редуцированный первый палец, II — V — хорошо развитые пальцы Пояс передних конечностей, или плечевой пояс, у земноводных, как и у акуловых рыб, лежит в толще мускулатуры туловища, связывающей его с осевым скелетом. Из верхней (дорзальной) лопаточной части первичного пояса образуется лопатка (scapula; рис. 61, 10); его самая верхняя часть остается хрящевой в виде широкого надлопаточного хряща (cartilago suprascapularis, рис. 61, 11). На передне-наружной поверхности надлопаточного хряща у некоторых бесхвостых амфибий имеется небольшое окостенение — остаток клейтрума рыбообразных предков. Окостеневшая коракоидная часть пояса превратилась в мощную коракоидную кость (coracoideum; рис. 61,12), вместе с лопаткой образующую суставную впадину для головки плеча (рис. 61, 13). Кпереди от коракоида за небольшим отверстием лежит хрящевой прокоракоид (cartilago procoracoidea; рис. 61, 14), на который налегает тоненькая покровная кость — ключица (clavicula; рис. 61, 15). Неокостеневшие хрящевые внутренние концы коракоидов и прокоракоидов правой и левой сторон сливаются вместе по средней линии. Позади коракоидов располагается костная грудина (sternum, рис. 62, 16) с хрящевым задним концом. Впереди от прокоракоидов выдается предгрудинник (praesternum; рис. 61, 17) также с хрящевым концом. В поясе передних конечностей хвостатых земноводных заметно больше хряща, а окостенения имеют меньшие размеры; ключицы часто не развиваются. Плечевой пояс служит опорой для передних конечностей и местом прикрепления управляющих ими мышц. Грудная клетка у земноводных не развивается: грудина не сочленяется с ребрами. 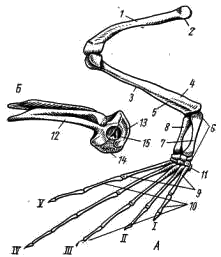 Рис. 62. Задняя конечность (А) и тазовый пояс (Б) лягушки сбоку: 1 — бедренная кость, 2 — головка бедра, 3 — голень, 4 — большая берцовая кость, 5 — малая берцовая кость, 6 — предплюсна, 7 — tibiale, 8 — fibulare, 9 — плюсна, 10 — фаланги пальцев, 11 — рудимент VI пальца, 12 — подвздошная кость, 13 — седалищная кость, 14 — лобковый хрящ, 15 — вертлужная впадина, I — V — пальцы Задняя конечность имеет удлиненную трубчатую кость — бедро (femur; рис. 62, 1), проксимальная часть которой заканчивается головкой (рис. 62, 2), входящей в вертлужную впадину (рис. 62, 15) тазового пояса. Большая берцовая (tibia; рис. 62, 4) и малая берцовая (fibula, рис. 62, 5) кости бесхвостых земноводных сливаются в единую кость голени (crus, рис. 62, 3); у хвостатых земноводных они сохраняются разделенными. Проксимальный ряд костей предплюсны (tarsus, рис. 62, 6) бесхвостых земноводных состоит из двух удлиненных костей, образующих добавочный рычаг конечности. Внутренняя из них называется tibiale (astragalus; рис. 62, 7; примыкает к большеберцовому краю голени), наружная — fibulare (calcaneus, рис. 62, 8). Между голенью и этими костями образуется голеностопный сустав. От дистального ряда костей предплюсны у земноводных сохраняются лишь 2 — 3 маленькие косточки. Плюсна (metatarsus; рис. 62, 9) образована пятью длинными косточками, к которым причленяются фаланги пальцев (phalanges digitorum; рис. 62, 10). Самый длинный палец у лягушек — IV. Сбоку от I (внутреннего) пальца располагается маленький рудимент VI («предпервого») пальца (praehallus; рис. 62, 11). Пояс задних конечностей, или тазовый, у земноводных, как и у всех наземных позвоночных, состоит из трех парных элементов; причем все они вместе образуют суставную вертлужную впадину (acetabulum; рис. 62, 15) для соединения с головкой бедра. Длинные, направленные вперед подвздошные кости (ilium; рис. 62, 12) своими концами причленяются к поперечным отросткам крестцового позвонка (см. рис. 54). Нижняя часть тазового пояса у земноводных не окостеневает и представлена лобковым хрящом (cartilago pubis, рис. 62, 14). Позади него лежат парные седалищные кости (ischium; рис. 62, 13). У хвостатых земноводных, по сравнению с бесхвостыми, в тазовом поясе много больше хряща, а сформировавшиеся кости малы. ЗАКЛЮЧЕНИЕ Земноводные (Amphibia) — первый класс наземных позвоночных животных. Однако у представителей класса еще сохраняется постоянная связь с водой. Эта двойственность отчетливо проявляется в особенностях эмбрионального и постэмбрионального развития. Яйцеклетки (икра) могут развиваться только в воде (или в редких случаях в предельно влажной среде). Из яйца выходит личинка — головастик, имеющая отчетливо выраженные признаки типичного водного животного: у него есть жабры и поддерживающие их жаберные дуги, сердце двухкамерное, один круг кровообращения, парные конечности наземного типа отсутствуют, основной орган движения — мощный хвостовой плавник, развиты органы боковой линии и т. п. Во время метаморфоза (превращения) личинка теряет часть признаков, свойственных водным животным, и приобретает особенности, обеспечивающие переход к наземному (вернее, наземно-водному) образу жизни: появляются парные конечности наземного типа, развиваются легкие, редуцируются жабры и поддерживающий их скелетный аппарат, перестраивается кровеносная система — образуются два неполностью разделенных круга кровообращения и т. п. Двойственность организации как приспособление к наземно-водному образу жизни хорошо выражена и у взрослых особей. Наземный образ жизни обеспечивается рядом особенностей строения: большая дифференцировка позвоночного столба на отделы и более прочное соединение тел позвонков друг с другом (замена амфицельных позвонков процельными или опистоцельными); появление парных конечностей наземного типа; усложнение строения и большая прочность поясов конечностей (при этом уже устанавливается достаточно прочное соединение тазового пояса с осевым скелетом); сильная редукция ме-тамерной мускулатуры и ее замена достаточно мощным сложным комплексом мускулов; появление век (защита глаз от механических повреждений, предотвращение подсыхания роговицы и т. п.); образование полости среднего уха с барабанной перепонкой и слуховой косточкой — стремечком (обеспечение возможности слуха в воздушной среде). Большую роль сыграло исчезновение жабр и развитие легких, гортани и хоан, создающих возможность воздушного дыхания; возникновение двух кругов кровообращения; большая дифференцировка пищеварительной системы (большие энергетические затраты при поддержании тела в воздушной среде) и т. п. Общее удлинение задних конечностей, обособление в них дополнительного рычага (благодаря резкому удлинению двух проксимальных косточек предплюсны) и возможность сильного перегиба посредине туловища в месте сочленения ветвей подвздошных костей с поперечными отростками крестцовых позвонков — приспособления к движению прыжками у бесхвостых земноводных. У ползающих хвостатых земноводных этих особенностей нет. Слияние в единое целое двух костей предплечья и двух костей голени связано с резким уменьшением необходимости вращательных движений стопы и кисти при перемещении прыжками. У хвостатых земноводных как предплечье, так и голень состоят из двух самостоятельных элементов, обеспечивая необходимые при ползании вращательные движения кисти и стопы. «Водные» черты строения проявляются в целом ряде особенностей: относительно слабое развитие окостенений скелета, обилие слизистых желез в коже (покрывающая кожу слизь уменьшает трение при движении в воде, предотвращает проникновение в кожу бактерий и грибков и т. п.), сохранение хвоста, часто уплощенного с боков и окаймленного кожистой складкой (тритоны и другие хвостатые земноводные), большое сходство мочеполовой системы с большинством групп рыб, характерное для подавляющего большинства видов земноводных наружное оплодотворение и т. д. При относительно небольшой поверхности легких земноводных необходимы достаточно мощные дополнительные органы дыхания. Таким органом становится всегда влажная (благодаря обилию слизистых желез), легко проницаемая для влаги и газов кожа и отчасти слизистая оболочка ротовой полости. У прудовой лягушки в активном состоянии легкие поглощают до 50% потребляемого организмом кислорода и выделяют только около 14% углекислого газа; путем кожного дыхания поглощается до 50% кислорода и выделяется до 86% углекислого газа. У ведущей более наземную жизнь травяной лягушки при легочном дыхании поступает до 67% кислорода и выделяется до 26% углекислого газа, а путем кожного дыхания поглощается 33% кислорода и выделяется 74% углекислого газа. При возрастании уровня метаболизма (усиление общей активности и всех процессов обмена при повышении температуры среды) удельная роль легких в обеспечении организма кислородом заметно возрастает. Понижение температуры среды вызывает снижение уровня метаболизма. При этом кожное дыхание почти в полной мере обеспечивает и насыщение организма кислородом и выделение углекислого газа, а относительное значение легких в дыхании резко снижается. Эта двойственность характера дыхания объясняется не только недостаточным развитием поверхности легких и несовершенством легочной вентиляции («заглатывание» воздуха при отсутствии грудной клетки); она необходима при земноводном образе жизни представителей этого класса. Именно эта двойственность органов дыхания обеспечивает земноводным возможность длительного пребывания в воде (вплоть до зимовки на дне водоема многих видов бесхвостых, когда при резком снижении уровня метаболизма кожное дыхание полностью обеспечивает все потребности организма в кислороде и выделении углекислого газа). Использование кожи для дыхания возможно лишь тогда, когда она легко проницаема для влаги и газов. Но такая кожа не может предохранять организм от больших потерь воды (высыхания). Поэтому практически почти все виды земноводных населяют лишь сырые, влажные участки, где организм теряет меньше влаги и всегда может восполнить ее потерю. У относительно мало связанных с водой жаб (зимуют на суше, в водоемы уходят только для икрометания) кожа уплотнена; это снижает возможности кожного дыхания, что компенсируется увеличением внутренней поверхности легких. Однако и у них, несмотря на уплотнение кожи, за ночной период охоты организм теряет до 15—30% воды. Некоторому уменьшению влагопотерь (при сохранении проницаемости кожи) у земноводных помогает слизь, покрывающая кожу. Обширные подкожные лимфатические полости служат резервуарами запасной воды. Уменьшаются влагопотери и благодаря обратному всасыванию воды в мочевом пузыре, заднем отделе кишечника и в клоаке. Очень резко уменьшаются влагопотери благодаря приспособительным особенностям поведения: повышенную активность земноводные проявляют только в часы максимальной влажности воздуха (в ясную погоду — в сумерках, а также ночью), на отдых они уходят в норки, где за счет почвенной влаги поддерживается высокая влажность. Двойственность органов дыхания делает невозможным полное разделение большого и малого кругов кровообращения. Но специфические особенности в строении сердца и отходящих от него кровеносных стволов обеспечивают некоторое разобщение кровотока несмотря на то, что в сердце земноводных только один желудочек, а в верхних полых венах есть примесь артериальной крови. Развитие мускульных выростов стенок желудочка уменьшает перемешивание крови, а от-хождение артериального конуса от правой (более венозной) стороны желудочка и детали его внутреннего строения (последовательность отхождения артериальных дуг, устройство спирального клапана) позволяют направлять более венозную кровь в кожу и легкие, более артериальную — к головному мозгу и органам чувств. Большая, по сравнению с рыбами, дифференцировка пищеварительного тракта приводит к некоторому повышению интенсивности использования пищи. Однако скорость пищеварения у земноводных невелика и зависит от температуры окружающей среды. Пищевые связи довольно простые; круг используемых кормов невелик (только животные относительно мелких размеров). Земноводные, как и рыбы, характеризуются непостоянством температуры тела (пойкилотермность): у земноводных она обычно лишь на 0,5—1°С выше температуры окружающей среды. Лишь в период наивысшей активности (преследование добычи, уход от опасности) температура тела может превышать температуру среды на 5—7°С. Пойкилотермность вызывает у земноводных умеренных и северных широт резко выраженную сезонную смену активности: при снижении температуры воздуха до +5 - +8°С все земноводные уходят в укрытия (часть видов лягушек — в ямы на дне водоемов; большинство видов бесхвостых и хвостатых земноводных скрываются в норах грызунов, прогнивших корнях деревьев, кучах мха и т.п.) и впадают в состояние оцепенения. С этим связано и географическое распространение земноводных: большинство видов этих животных свойственны тропической зоне. В тропиках, при относительно устойчивых в течение года температурных условиях, у ряда видов земноводных состояние оцепенения вызывается резким снижением влажности воздуха («спячка» в сухой период года). Очень большая зависимость земноводных от влажности и температуры среды сказывается в том, что погодные условия (в наших широтах — сильные засухи летом, сильные морозы без снега зимой) часто служат основной причиной смертности и определяют резкие колебания численности земноводных по годам. Дополнительная литература |