Карташев Наумов Практикум по зоологии. "Практикум по зоологии позвоночных". Подтип бесчерепные acrania тема строение ланцетника систематическое положение объекта
 Скачать 5.56 Mb. Скачать 5.56 Mb.
|

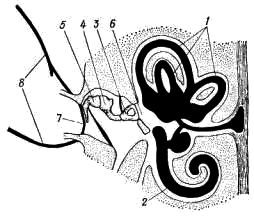 Рис. 169. Схема строения уха млекопитающих: 1 — полукружные каналы, 2 — улитка, 3 — стремечко, 4 — наковальня, 5 — молоточек, 6 — овальное окно, 7 — барабанная перепонка, 8 — наружное ухо Для упрощения описания зубов употребляют так называемые зубные формулы: в числителе указывают число зубов одной половины верхней челюсти, в знаменателе — нижней. Сокращенно обозначают: резцы — i, клыки — с, предкоренные — р, коренные — m. Например, зубная формула лисицы имеет следующий вид: i 3/3 c 1/1 p 4/3 m 2/3 = 42 Конечности и их пояса. Пояс передней конечности лисицы состоит только из лопатки (scapula; рис. 170, 1). Треугольная по форме, широкая и тонкая лопатка на латеральной поверхности несет высокий гребень (spina scapulae; рис. 170, 2), заканчивающийся акромиальным отростком (processus acromion; рис. 170, 3). В дистальном отделе лопатки располагается суставная ямка (рис. 170, 4), в которую входит головка плечевой кости. Около суставной ямки виден коракоидный отросток (processus coracoideus; рис. 170, 5), представляющий собой редуцированный и приросший к лопатке коракоид. Ключица у лисицы отсутствует, как и у большинства хищных (а также копытных, хоботных и китообразных). У большинства сумчатых, насекомоядных, рукокрылых, приматов, грызунов, зайцеобразных ключицы (clavicula) развиты хорошо. Передняя конечность состоит из трех отделов: плеча, предплечья и кисти. В плечевом отделе всего одна кость — плечевая (humerus; рис. 170, 6). Ее проксимальный отдел заканчивается округлой головкой, входящей в суставную ямку лопатки, а дистальный отдел — блоковидным выступом, сочленяющимся с предплечьем. Предплечье образовано локтевой костью (ulna; рис. 170, 7) и несколько более толстой лучевой (radius; рис. 170, 8). Локтевая кость имеет в проксимальном отделе крупный локтевой отросток. Кисть разделяется на проксимальный отдел — кости запястья (carpus; рис. 170, 9), промежуточный — кости пясти (metacarpus; рис. 170, 10) и дистальный — фаланги пальцев (phalanges digitorum; рис. 170, 11). Тазовый пояс состоит из двух безымянных костей (innominatum; рис. 171). Каждая из них образована срастанием костей; подвздошной (ilium; рис. 171, 1), седалищной (ischium; рис. 171, 2) и лобковой (pubis; рис. 171, 3). В месте схождения этих костей находится вертлужная впадина (acetabulum; рис. 171, 4), служащая для сочленил с бедренной костью. Таз млекопитающих закрытый: лобковая и седалищная кости правой и левой сторон срастаются друг с другом по средней линии. 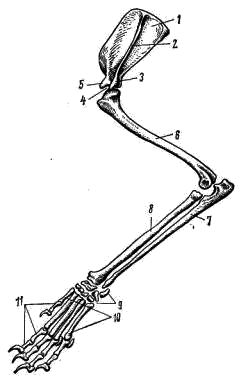 Рис. 170. Плечевой пояс и передняя конечность лисицы: 1 — лопатка, 2 — гребень лопатки, 3 — акромиальный отросток, 4 — суставная ямка, 5 — коракоидный отросток, 6 — плечевая кость, 7 — локтевая кость, 8 — лучевая кость, 9 — запястье, 10 — пясть, 11 — фаланги пальцев 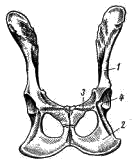 Рис. 171. Тазовый пояс лисицы (вид снизу): 1 — подвздошная кость, 2 — седалищная кость, 3 — лобковая кость, 4 — вертлужная впадина В задней конечности имеется три отдела: бедро, голень и стопа. Бедренный отдел содержит одну кость — бедро (femur; рис. 172, 1). Проксимальный отдел бедра имеет головку, которая входитввертлужную впадину таза. Бедро сочленяется с голенью коленным суставом, на передней поверхности которого расположена небольшая округлая косточка — коленная чашечка (patella; рис. 172, 2). Голень имеет крупную большую берцовую кость (tibia; рис. 172, 3) и тонкую малую берцовую кость (fibula; рис. 172, 4). Стопа образована предплюсной (tarsus; рис. 172, 5), плюсной (metatarsus; рис. 172, 6) и фалангами пальцев (phalanges digitorum; рис. 172, 7). Предплюсна в проксимальном отделе содержит две косточки: внутреннюю — таранную (astragalus; рис. 172, 8) и наружную — пяточную (calcaneus; рис. 172, 9) с направленным назад пяточным выступом. В отличие от пресмыкающихся и птиц у млекопитающих сустав, обеспечивающий подвижность стопы, располагается между костями голени и проксимальными костями предплюсны; такой сустав называется голеностопным. 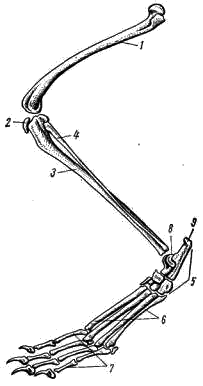 Рис. 172. Задняя конечность лисицы: 1 — бедренная кость, 2 — коленная чашечка, 3 — большая берцовая кость, 4 — малая берцовая кость, 5 — предплюсна, 6 — плюсна, 7 — фаланги пальцев, 8 — таранная кость, 9 — пяточная кость ЗАКЛЮЧЕНИЕ Предками млекопитающих были примитивные пресмыкающиеся, еще имевшие ряд общих признаков с земноводным. В связи с этим современные млекопитающие имеют определенное сходство в строении не только с пресмыкающимися, но и с земноводными. Конечности млекопитающих, как и земноводных, имеют голеностопное и предплечно-запястное сочленения, череп обладает широким основанием (платибазальный тип), в коже находятся многочисленные железы. С пресмыкающимися млекопитающих сближают общие черты в строении скуловой дуги (синапсидный тип, свойственный звероящерам: имеется одна скуловая дуга, образованная элементами как нижней — скуловая кость, так и верхней — чешуйчатая кость — височных дуг черепа диапсидного типа). Некоторые современные примитивные млекопитающие обнаруживают и другие черты сходства с пресмыкающимися: так, плечевой пояс однопроходных имеет надгрудинник, коракоид и прокоракоид; эти животные имеют клоаку и откладывают яйца. На фоне сходства с земноводными и пресмыкающимися млекопитающие характеризуются высокой степенью организации, что выражается, прежде всего, в прогрессивном развитии нервной системы. В головном мозгу крупных размеров достигает передний мозг (полушария), покрывающий сверху большую часть остальных отделов. Полушария могут достигать половины массы всего головного мозга (крот, кролик) и еще больше (до 4/5 массы всего мозга у человека). Строение переднего мозга млекопитающих весьма специфично. Если у рыб, земноводных, пресмыкающихся и птиц основную массу переднего мозга составляют полосатые тела, занимающие нижние участки полушарий, и крыша мозга (свод) остается тонкой, то у млекопитающих наиболее сильное развитие получает именно свод. Свод у млекопитающих дифференцируется на первичный (archipallium) и вторичный (neopallium); преимущественное развитие имеет последний. Вторичный свод представляет собой скопление нервных клеток (серое вещество мозга), расположенных по периферии полушарий; так образуется кора головного мозга. Благодаря этому увеличивается количество нервных клеток в мозгу; кроме того, поверхностное расположение обеспечивает лучшее их кровоснабжение. Толщина серого вещества — коры полушарии у различных млекопитающих примерно одинакова, и увеличение числа нервных клеток может достигаться лишь увеличением поверхности коры за счет образования борозд и складок. Поэтому полушария переднего мозга высших млекопитающих (особенно копытных, хищных, приматов) имеют сложную систему борозд и извилин. Для различных систематических групп млекопитающих характерен свой тип складчатости коры. Кора полушарий содержит ассоциативные центры, связанные с высшей психической деятельностью, а также высшие чувствующие (зрительные, слуховые и осязательные) и двигательные центры. В связи с высокой подвижностью млекопитающих крупных размеров достигает мозжечок. Усложняется его внутреннее строение, усиливаются связи с двигательными центрами коры полушарий переднего мозга. Прогрессивно развитая нервная система млекопитающих определяет значительно более сложное поведение, чем у представителей других классов позвоночных, что, естественно, дает им дополнительные шансы в борьбе за жизнь. Благодаря совершенной сложнорефлекторной деятельности млекопитающие обладают несравненно более высокой способностью приспосабливаться к окружающей среде посредством изменения своего поведения, чем другие позвоночные. Скрытный, часто ночной или сумеречный, образ жизни большинства млекопитающих ограничивает возможность применения зрения, компенсируя его сильным развитием обоняния и слуха. Обоняние у млекопитающих более тонкое, чем у других позвоночных. Орган обоняния располагается в верхнезадней части носовой полости. Сложная система обонятельных раковин резко увеличивает площадь, занятую обонятельным эпителием (кроме того, сложная конфигурация обонятельной полости способствует очищению, согреванию и увлажнению вдыхаемого воздуха). Обоняние имеет важное значение не только при межвидовых (поиск пищи, распознавание врагов и т. д.), но и во внутривидовых отношениях (в период размножения, при охране индивидуальных участков и т. д.). В последнем случае важное значение имеет то, что многие из кожных желез выделяют пахучий секрет, который придает млекопитающим не только видовой, но и индивидуальный запах. Орган слуха млекопитающих, по сравнению с другими позвоночными, имеет более сложное строение. Появляется хорошо развитое наружное ухо, представленное ушной раковиной и наружным слуховым проходом. Подвижная ушная раковина концентрирует и направляет в наружный слуховой проход звуковые волны, которые колеблют барабанную перепонку. В среднем ухе млекопитающих расположены три подвижно сочлененные между собой слуховые косточки. В дополнение к стремечку (stapes), имевшемуся у земноводных, пресмыкающихся и птиц, у млекопитающих появляются еще молоточек (malleus) и наковальня (incus). Молоточек образуется из сочленовной кости, а наковальня — из квадратной. Колебания барабанной перепонки передаются через слуховые косточки перилимфе внутреннего уха благодаря давлению стремечка на эластичную перепонку овального окна внутреннего уха. Рычажная система слуховых косточек позволяет млекопитающим улавливать слабые звуки. Улитка внутреннего уха достигает высокого развития, и в ней обособляется сложно устроенный кортиев орган, который собственно и служит воспринимающим отделом органа слуха. Орган зрения млекопитающих, как правило, не является ведущим рецептором. Аккомодация глаза достигается только путем изменения кривизны хрусталика расположенной по его периферии ресничной мышцей. Прогрессивно развитие относительно большого поля бинокулярного зрения, позволяющего фокусировать оба глаза одновременно на одном предмете. Как и у птиц, у млекопитающих происходит интенсификация всех жизненных процессов. Это обеспечивается рядом приспособлений. Ткань легких млекопитающих состоит из многочисленных легочных пузырьков — альвеол, способствующих значительному увеличению поверхности соприкосновения респираторного эпителия с воздухом. Например, легкие лошади насчитывают примерно 5 млрд. альвеол с общей поверхностью в 500 м2. Такая большая дыхательная поверхность легких увеличивает возможность газообмена, соответственно повышая общую жизнедеятельность организма. Увеличению газообмена способствуют также высокая степень насыщения кровеносными сосудами стенок альвеол и интенсивный ток крови в этих сосудах (значительно более интенсивный, чем у пресмыкающихся). Акт дыхания у млекопитающих происходит не только путем расширения и сужения грудной клетки, как у других амниот, но и одновременным опусканием и подъемом диафрагмы. Система органов кровообращения у млекопитающих также имеет прогрессивные черты. Для форменных элементов крови, обеспечивающих ее дыхательную функцию, — эритроцитов, характерны отсутствие ядер, небольшие размеры и высокая численность их в крови. Маленькие эритроциты, при одинаковой форме с большими, имеют относительно большую поверхность при той же массе вещества. Следовательно, при уменьшении размеров эритроцитов возрастает их общая дыхательная поверхность. Отсутствие ядер в эритроцитах влечет за собой понижение обмена веществ этих клеток, в связи с чем они сами меньше потребляют кислорода и больше отдают его тканям. Несмешанное кровообращение обеспечивает лучшее снабжение тканей тела кислородом и питательными веществами. У млекопитающих такой тип кровообращения достигается, как и у птиц, полным разделением сердца на правую (венозную) и левую (артериальную) половины и редукцией одной из дуг аорты. В отличие от птиц млекопитающие имеют не правую, а левую дугу аорты. Прогрессивное развитие пищеварительной системы млекопитающих (дифференциация зубной системы и пищеварительного тракта) позволило им, с одной стороны, использовать в качестве пищи самые разнообразные растительные и животные объекты, а с другой — значительно интенсифицировать процесс пищеварения. Разнокачественность зубов (гетеродонтность) возникла в результате специализации различных групп зубов для схватывания добычи, разгрызания, пережевывания. Измельчение пищи во время жевания облегчает и убыстряет пищеварительный процесс. Сложная дифференцировка пищеварительного тракта и деятельность многочисленных пищеварительных желез способствуют интенсивному перевариванию пищи с помощью разнообразных ферментов. Большое значение в пищеварении имеет симбиотическая бактериальная флора кишечника, особенно сильно развитая у растительноядных форм. Перечисленные приспособления обусловили высокий уровень обмена веществ млекопитающих. Интенсивный обмен веществ и развившаяся система терморегуляции позволяют этим животным поддерживать температуру своего тела постоянной, что делает их в известной степени независимыми от температуры окружающей среды. В поддержании постоянной температуры тела важное значение имеет волосяной покров, обеспечивающий термоизоляцию и отчасти терморегуляцию. Важную роль в теплозащите организма может играть и подкожный слой жира. Потовые железы, имеющие важное значение как дополнительные органы выделения, могут служить у млекопитающих и целям терморегуляции, смачивая поверхность тела потом, который, испаряясь, охлаждает организм. Млекопитающие в огромном своем большинстве значительно более подвижны и активны, чем пресмыкающиеся. В связи с этим у них прогрессивное развитие получают соответствующие элементы скелета и мышечной системы. Постановка конечностей млекопитающих отличается более совершенным типом: они не растопырены в стороны, и тело не провисает между ними, как у пресмыкающихся. В связи с этим упрощается строение плечевого пояса. Основной толчок при передвижении осуществляется задними конечностями, поэтому у большинства млекопитающих их скелет и мускулатура развиты относительно сильнее. В зависимости от экологических особенностей строение конечностей, особенно их дистальных отделов, у разных видов млекопитающих сильно варьирует. S-образная изогнутость позвоночника в вертикальной плоскости обеспечивает достаточно успешное выполнение им опорной функции и амортизацию при беге, прыжках и т. п. В последнем случае важное значение имеют хрящевые мениски, расположенные между позвонками. Значительно более прогрессивны по сравнению с другими позвоночными особенности размножения млекопитающих. Формирование зародыша происходит в общем сходным образом с пресмыкающимися и птицами. Развитие же зародышевых оболочек млекопитающих весьма специфично. Желточный мешок зародыша, полость которого не имеет питательных веществ, исчезает рано. На ранних стадиях у зародыша появляются амнион и аллантоис, которые достигают сильного развития. Сероза срастается с внешней стенкой аллантоиса и дает многочисленные отростки, врастающие в стенку матки — образуется так называемая плацента, характерная для большинства млекопитающих. Плацента — приспособление, с помощью которого зародыш может питаться и дышать за счет материнского организма. Кровеносные сосуды зародыша и матери настолько тесно соприкасаются друг с другом, что обеспечивается поступление питательных веществ и кислорода из крови матери в кровь зародыша и отдача в обратном направлении продуктов обмена. После рождения детеныши млекопитающих вскармливаются молоком матери, продуцируемым специфичными млечными железами — видоизмененными потовыми железами кожи. Длительное развитие детенышей в утробе матери (или в сумке) и вскармливание их после рождения молоком способствуют лучшему выживанию молодняка. Отмеченные выше прогрессивные особенности млекопитающих обеспечили им возможность приспособления к различным условиям существования и позволили широко расселиться по земному шару, образовав наземные, подземные, водные и воздушные формы. Еще более разнообразные, чем у птиц, приспособления позволяют млекопитающим переживать неблагоприятные сезоны года. В умеренных и северных широтах при наступлении зимы некоторые млекопитающие мигрируют на юг, иногда покрывая расстояния в несколько тысяч километров (например, северные олени,некоторые летучие мыши, китообразные и др.). Другие млекопитающие совершают лишь небольшие кочевки, переходя в более кормные участки, или же ведут оседлый образ жизни. Для оседлых видов обычно характерны сезонные изменения суточной активности, структуры волосяного покрова и его теплоизолирующих свойств, набора кормов, характера обмена веществ и т. п. Для некоторых видов свойственно запасание корма (бурундук, бобр, многие полевки и др.). Некоторые млекопитающие переживают неблагоприятные сезоны, впадая в спячку (ежи, сурки, суслики, оседлые виды летучих мышей и др.). Дополнительная литература Астанин Л. П. Органы тела млекопитающих и их работа. М., 1958. Барабаш-Никифоров И. И., Формозов А. Н. Териология. М., 1963. Бобринский Н. А., Кузнецов Б. А., Кузякин А. П. Определитель млекопитающих. М., 1965. Громов И. М. и др. Млекопитающие фауны СССР, ч. 1, 2, М.—Л., 1963. Огнев С. И. Экология млекопитающих. М, 1951. Соколов В. Е. Систематика млекопитающих, ч. 1, 1973; ч. 2, 1977; ч. 3, 1979. Шмальгаузен И. И. Основы сравнительной анатомии. М., 1947. ТЕМА 21. ОПРЕДЕЛЕНИЕ МЛЕКОПИТАЮЩИХ МАТЕРИАЛ И ОБОРУДОВАНИЕ Набор коллекционного материала (тушки млекопитающих с очищенными черепами и этикетками с указанием основных промеров). На одного-двух студентов необходимы: 1. Ручная лупа 2—4 X. 2. Штангенциркуль. ЗАДАНИЕ 1. Определить по одному представителю (по черепам) всех имеющихся в наборе отрядов млекопитающих; кратко записать основные «ключевые» признаки каждого отряда. 2. По указанию преподавателя провести определение до вида, последовательно устанавливая отряд, семейство, род и вид. 3. Обратить внимание на связь особенностей внешнего облика с образом жизни (приспособления к водному, древесному, роющему образу жизни и т. д.). Распределить определенные виды по их образу жизни. Прежде чем приступить к определению, необходимо ознакомиться с главными стандартными промерами тела и черепа, принятыми в систематике млекопитающих. 1. Длина тела у животных не крупнее зайца или лисицы измеряется со стороны брюха от конца морды до заднепроходного отверстия; у более крупных зверей — по спине от конца морды до основания хвоста. 2. Длина хвоста — расстояние от основания хвоста до его конца без концевых волос. 3. Длина ступни — расстояние от задней части пятки до конца самого длинного пальца без когтя. 4. Длина (или высота) уха — расстояние от нижнего края ушной раковины до вершины уха без концевых волос. 5. Наибольшая длина черепа (рис. 173, а) — расстояние от наиболее выступающей вперед точки верхних резцов до наиболее выдающейся назад части черепа. |