реферат по теме Костистые рыбы. Костистые рыбы. Развитие костистых рыб
 Скачать 1.23 Mb. Скачать 1.23 Mb.
|

РАЗВИТИЕ КОСТИСТЫХ РЫБЭМБРИОНАЛЬНЫЙ ПЕРИОДПОДПЕРИОД - РАЗВИТИЕ В ОБОЛОЧКЕ I. Этап. Активация яйца и образование бластодискаПосле соединения гамет начинается активация яйца, что заметно по выделению содержимого кортикальных альвеол (гранул) и образованию перивителлинового пространства. Цитоплазма стягивается на анимальный полюс и образует бугорок – бластодиск (рис. VIII.1, А). На протяжении этапа завершается второе деление мейоза, сближаются мужской и женский пронуклеусы и вскоре появляется веретено первого деления дробления. На протяжении этапа происходит затвердевание, т. е. увеличение прочности оболочки (Зотин, 1961). этап. ДроблениеДроблением называется серия митотических делений ядра и цитоплазмы зиготы, не сопровождающихся клеточным ростом. Это приводит к образованию многоклеточного зародыша. Первая борозда дробления обычно проходит через центр бластодиска в меридиональном направлении и делит бластодиск на два, обычно разных по величине бластомера(рис. VIII.1, Б). Вторая борозда дробления, также меридиональная, проходит перпендикулярно первой борозде, и в результате образуются четыре бластомера (рис. VIII.1, В). Вторая борозда представляет собой также плоскость билатеральной симметрии зародыша. Две борозды третьего деления дробления проходят параллельно одна другой через центры лежащих в два ряда бластомеров, образуя 8 клеток (рис. VIII.1, Г). Две последующие борозды четвертого деления приводят к образованию 16 бластомеров. Особенность первых четырех делений состоит в том, что деления клеток (цитокинезы) не завершаются и плазмалемма в нижней части бластомеров не образуется. Все клетки соединены в нижней части общим цитоплазматическим слоем, образуя синцитий.  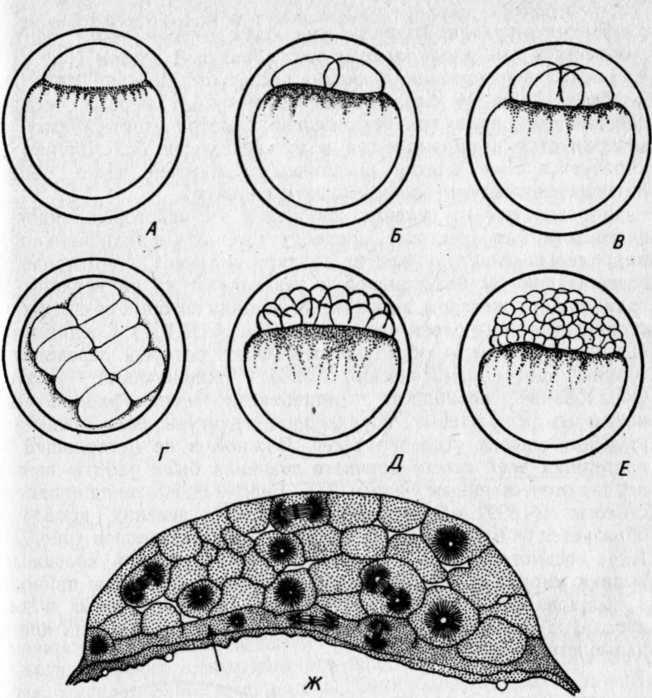 Рис. VIII.1. Ранние этапы развития карпа (А–Е, ориг.) и щуки (Ж, по Бергу, 1899). А – 1 этап – активация яйца и образование плазменного бугорка (температура 26°, возраст 40– 50 мин). II этап – дробление: Б – 2 бластомера (около 1 ч), В – 4 бластомера (1 ч 20 мин), Г – 8 бластомеров (1 ч 40 мин), Д – 32 бластомера (2 ч 20 мин), Е – 128 бластомеров (около 3 ч). Ж – яйцо щуки в конце дробления (стрелкой отмечен перибласт) Борозда пятого деления дробления (у некоторых видов четвертого или шестого) проходит уже не меридионально, а параллельно экватору желтка (латитудинально) и это приводит к появлению 32 бластомеров и разделению их на верхние, не граничащие с желтком, и нижние, основания которых по-прежнему соединены общим слоем цитоплазмы (рис. VIII.1, Д). Соединенные нижние бластомеры образуют перибласт, который переходит в тонкий цитоплазматический слой, окружающий желток. Образование перибласта (парабласта) – специфичной для костистых (и хрящевых) провизорной структуры, было предметом изучения многих исследователей. Важной и не потерявшей до настоящих дней своего научного значения была работа, выполненная отечественным ученым Л. С. Бергом еще в конце прошлого столетия (1899). В результате шестого деления дробления образуется 64 бластомера, седьмого – 128 бластомеров (рис. VIII.1, Е), восьмого – 256. Стадии 64–256 бластомеров иногда называют «морулой». Последующие деления бластомеров приводя к образованию большого числа клеток, расположенных в виде купола на перибласте (рис. VIII.1, Ж). Совокупность этих клеток называется бластодермой. Форма бластомеров на протяжении клеточного цикла не остается неизменной, а постоянно меняется. Перед очередным делением бластомеры становятся выше и соприкасаются между собой. После деления клетки слегка уплощаются и контакты между ними ослабевают (рис. VIII.2, А). Эти свойства бластомеров связаны с изменением состояния их мембран. Появление борозд дробления на поверхности цитоплазмы происходит после метафазы и приходится на ана-, тело- или интерфазу.  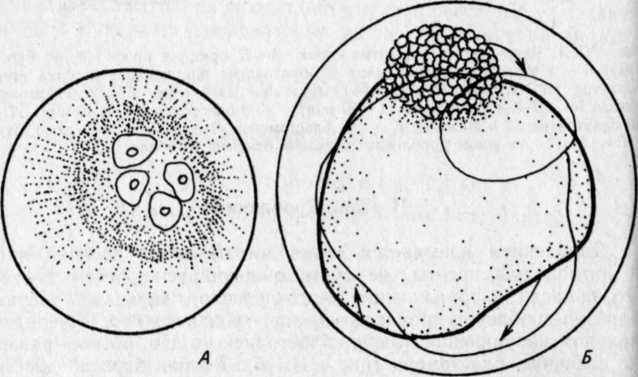 Рис. VIII.2. Ранние стадии развития канального сома (ориг.). А – стадия 4 бластомеров во время исчезновения между ними контактов. Б – цитоплазматическая моторика на стадии ранней бластулы (стрелками отмечено изменение положения бластодермы и формы желточного мешка) Особенность первых делений дробления состоит в их строгой синхронности. Число синхронных делений различно у разных видов, обычно их 4–6. Для этих делений характерна максимальная скорость. Она определяется спецификой клеточного цикла, в котором отсутствует в интерфазе фаза G1 (табл. 3)1  1 Клеточный цикл включает интерфазу и митоз. Интерфаза начинается стадией пресинтеза (Gi), когда идут процессы функционировании генов и подготовительные процессы к синтезу ДНК: центральное место в интерфазе занимает фаза S – синтеза, ауторепродукция хромосом; в это время количество ДНК в клетке удваивается и вместо одной хромосомы появляются две хроматиды; завершает интерфазу фаза Gi, на которой начинается упаковка и спирализация хромосом, т. е. подготовка к будущему митозу. Митоз состоит из четырех фаз: про-, мета-, ана- и телофазы. Таблица 3  На 5–7-м циклах начинается десинхронизация дробления и последующие деления клеток происходят все более и более асинхронно. Десинхронизация клеточных делений начинается с возникновения «митотического градиента» или «волны митозов», распространяющейся от верхних бластомеров к нижним. Десинхронизация делений клеток свидетельствует о начавшейся дифференциации бластомеров, но четких внешних показателей этого процесса еще нет. Однако электронномикроскопические исследования выявили различия в распределении в верхних и нижних бластомерах белковых (желтка) включений. Одной из причин «волны митозов» может быть неравномерное распределение этих включений в бластомерах разных участков бластодермы. Другая причина может состоять в изменении межклеточных обменных процессов, о чем судят по уменьшению величины электрической связи между клетками. Несмотря на десинхронизацию делений, короткие митотические циклы, свойственные синхронным делениям, сохраняются еще на протяжении 5–7 делений. Таким образом, число быстрых делений составляет у разных видов рыб 11–13. Количество клеток на этом отрезке развития достигает нескольких тысяч: у лосося – двух, у вьюна – восьми (Ротт, 1980, 1987). У некоторых видов на этапе дробления начинает проявляться цитоплазматическая моторика, выражающаяся в прохождении по цитоплазме волны сокращения. Это приводит к изменению формы желточного мешка и покачиванию яйца в яйцевой оболочке, что способствует устранению вокруг него микрозоны с низким содержанием кислорода. этап. БластуляцияНа этапе бластуляции еще продолжается дробление бластомеров и впервые становится заметной дифференциация клеток. Поверхностные клетки бластодермы уплотняются и приобретают полигональную форму. Их боковые клеточные мембраны прилегают друг к другу, и между ними возникают контакты. В результате этого на поверхности бластодермы появляется эпителиальный пласт – перидерма2, характерная для костистых рыб (как, впрочем, и для костных ганоидов) структура (см. обзор Детлаф, 1982). Под перидермой находятся глубокие клетки, которые неплотно прилегают друг к другу, между ними имеются щели и полости, представляющие собой полость дробления – бластоцель(или 2 Перидерма на ранних стадиях развития костистых рыб выполняет функцию покровов. У одних рыб (лососевые) клетки ее отпадают перед вылуплением зародыша (Bouvet, 1976), у других – перидерма принимает участие в образовании провизорных структур у зародышей и предличинок (см. разд. VIII.2) первичную полость тела). У большинства костистых рыб четко выраженного бластоцеля, как у осетровых и двоякодышащих рыб, нет. Видимые иногда на гистологических срезах большие полости образуются при обезвоживании зародышей в результате разного сжатия клеточного материала и желтка и являются артефактом. В эмбриологии рыб условно выделяют раннюю, среднюю и позднюю бластулы. Для ранней бластулы характерен высокий купол клеток бластодермы, средней – небольшое его уплощение, поздней – значительное уплощение и впячивание в бластодерму верхней части желточного мешка. На бластуле у некоторых видов сильно выражена цитоплазматическая моторика (рис.VIII.2, Б). Она приводит к вращению икринки вокруг своей оси и перемешиванию перивителлиновой жидкости. Глубоким клеткам бластодермы на этапе бластулы уже свойственны слабые движения. Они выражаются в периодическом появлении у них коротких выступов лобоподий, которыми клетки касаются друг друга, не образуя между собой контакта и не совершая направленных движений. Поверхность желтка под бластодермой покрыта перибластом. За границей бластодермы на поверхности желтка перибласт переходит в тонкий цитоплазматический слой, окружающий желток. Значение перибласта состоит в переработке желтка и проведении питательных веществ в бластоцель, откуда они поступают в клетки. Этой цели служат ворсинки перибласта, направленные в бластоцель, и тяжи цитоплазмы перибласта, проникающие в желток. Для процесса бластуляции характерно удлинение клеточного цикла, а именно интерфазы за счет увеличения фазы S и появления фазы G1, отсутствовавшей на этапе дробления. В ядрах возникают ядрышки и активируется синтез РНК, в цитоплазме возрастает число клеточных органелл и белковых гранул. Перестройка клеточных циклов – важное событие в раннем эмбриогенезе рыб. Эта перестройка создает условия для морфогенетической функции ядер самого зародыша и означает переход от дробления к бластуляции. Относительно причин, вызывающих перестройку клеточных циклов, имеется несколько гипотез. Согласно одной из них сигналом к изменению циклов служит достижение клетками определенных величин ядерно-плазменных отношений. Вследствие отсутствия при дроблении роста клеток объем цитоплазмы в каждом бластомере после очередного деления уменьшается в два раза, в то время как содержание ДНК остается на прежнем уровне. Таким образом, ядерно-плазменное отношение все время возрастает пока не достигает величин, требующих перестройки клеточного цикла. По другой гипотезе перестройку клеточных циклов вызывает изменение отношения в бластомерах ионов K+ и Na+. В процессе развития ионы K+ поступают в бластомеры из желтка, поэтому количество K+ в них увеличивается. Содержание же Na+ не меняется. В результате отношение K+/Na+ при развитии возрастает. К моменту замедления клеточных делений, это отношение становится равным двум. Такое отношение отмечено у всех исследованных рыб, а также амфибий. Очевидно, связь между отношением K+/Na+=2 и перестройкой клеточного цикла является закономерной (Ротт, 1987). этап. ГаструляцияГаструляцией традиционно называют процесс разделения однородной бластодермы бластулы на два слоя клеток – зародышевые листки. Различают внутренний и наружный листки – соответственно энтодерму и эктодерму. Позднее между этими листками появляется третий листок – мезодерма. Гаструляция в разных систематических группах животных идет разными способами. Она осуществляется благодаря интенсивным морфогенетическим перемещениям клеток, и результат ее всегда один – образуется комплекс осевых зачатков, расположенных соответственно основному плану строения животного. Процесс гаструляции у костистых рыб, как показали исследования Балларда (Ballard, 1973, 1982), весьма своеобразен и отличается от этого процесса у прочих позвоночных. У рыб не происходит подворачивания бластодермы и, по существу, не образуются зародышевые листки, а происходит в закладывающемся теле зародыша рассортировка клеток согласно их детерминации. У костистых рыб гаструляция осуществляется двумя типами перемещения клеток бластодермы: обрастанием желтка, т. е, эпиболией, при которой клетки мигрируют в меридиональном направлении, и осевой конвергенцией, при этом клетки движутся в широтном направлении – к будущей оси зародыша. При переходе к гаструляции происходят существенные изменения в структуре и свойствах клеток бластодермы. В их цитоплазме возрастает число клеточных органелл и усложняется их строение. Это приводит к увеличению активности клеток, установлению между ними контактов и слипанию (адгезии) их. Короткие выступы глубоких клеток – лобоподии – превращаются в длинные отростки с веерообразными мембранами на концах – филоподии, благодаря которым и осуществляются контакты и перемещения клеток. В начале гаструляции происходит эвакуация глубоких клеток из центральной части бластодермы к ее периферии. При этом между перибластом и перидермой возникает полость, расположенная немного асимметрично по отношению к центру зародышевого диска. Обычно эту полость считают бластоцелем, однако Баллард полагает, что она не гомологична бластоцелю, и называет ее полостью желточного мешка, внутризародышевой3. Глубокие клетки, мигрировавшие из центра бластодермы, образуют по краю зародышевого диска утолщение – зародышевое кольцо. В дорсальном секторе бластодермы (будущем головном отделе зародыша) происходит концентрация глубоких клеток – зародышевый узелок, который вскоре удлиняется вследствие поступления нового клеточного материала и преобразуется в зародышевый щиток. Глубокие клетки бластодермы у одних рыб (треска Gadusmorhuamarisalbi) еще до миграции из центральной части бластодермы, у других (сайка Boreogadus saida) – в зародышевом кольце образуют два слоя клеток: верхний – эпибласт, прилегающий к нижней поверхности перидермы, и нижний – гипобласт, располагающийся на перибласте (Махотин, 1982). Эпиболия начинается с разрастания перибласта, находящегося под бластодермой на анимальном полюсе, называемого внутренним или базальным 3 Эта полость у многих видов рыб с донной икрой сохраняется довольно - долго, она служит для ориентации зародышей головой вверх, и называют ее перибластическим синусом (Соин, Черняев, 1961) (см. рис. VI.2). (Доронин и др., 1989). Краевая зона перибласта выступает за край бластодермы и постепенно распространяется в сторону вегетативного полюса. Одновременно перибласт слегка смещается в сторону будущей оси зародыша (Long, 1983). Распространяющийся по желтку перибласт называют наружным. Краевые клетки перидермы слипаются с перибластом, образуя край обрастания. Перибласт, разрастаясь, увлекает за собой перидерму. В начале и конце обрастания перибласт обычно распространяется впереди клеточного материала (Тринкаус, 1972). Перемещение глубоких клеток бластодермы происходит в составе зародышевого кольца. Достигнув вегетативного полюса, наружный перибласт смыкается, затем срастаются перидерма и края бластодермы, сначала окружая, потом замыкая остаточную часть желтка – желточную пробку(рис.VIII.З).  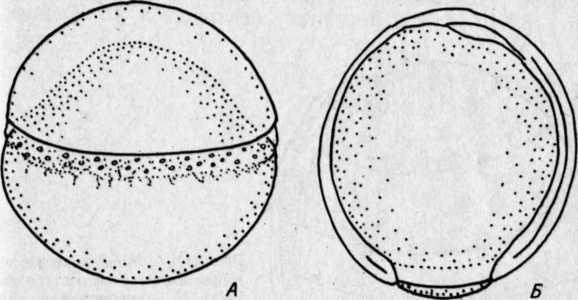 Рис. VIII.3. IV этап – гаструляции у карпа (ориг.). А – начало обрастания (7 ч), Б – завершение обрастания, заметны закладка тела зародыша и небольшая желточная пробка (9 ч) Желток, окруженный перибластом и клеточным материалом, называется желточным мешком. Его рассматривают как провизорный орган зародыша (Токин, 1987) и как первичную кишку (Бочаров, 1988). На протяжении эпиболии область зародыша, испытывающая цитоплазматическую моторику, становится все меньше, так как желток покрывается клеточным материалом. Осевая конвергенция. Перемещение глубоких клеток из бластодермы зародышевого диска происходит по радиусам, причем клетки, располагавшиеся в дорсальном секторе, мигрируют пучком непосредственно в зародышевый щиток. Клетки боковых и вентрального секторов мигрируют сначала в зародышевое кольцо по меридианам яйца, но при этом всегда уклоняются в дорсальную сторону (рис.VIII.4. А, Б). Достигнув кольца, они меняют путь миграции и начинают перемещаться к закладке зародыша, т. е. в дорсальном направлении, где и происходит их концентрация (рис.VIII.4. В). Эти перемещения глубоких клеток в широтном направлении к оси зародыша называются осевой конвергенцией. Как показали эксперименты, проведенные на зародышах форели, осевая конвергенция клеток гипобласта определяется соответствующими конвергентными перемещениями наружного перибласта, выполняющего важную роль в обеспечении билатеральной симметрии зародыша (Long, 1983). Клетки гипобласта активны и подвижны. Они часто располагаются двумя слоями. Нижний из них, будущая энтодерма, образован одним рядом рыхло расположенных клеток. Верхний слой, будущая мезодерма, представлен клетками, расположенными более плотно и в несколько рядов. Клетки гипобласта в зародышевом кольце мигрируют в сторону вегетативного полюса, уклоняясь к оси зародыша. Скорость их перемещений составляет у фундулуса 10–15 мкм/мин. При замыкании желточной пробки остаточная мезодермальная часть гипобласта не включается в туловище зародыша и образует в области пробки скопление клеток – ареолу. Эпибласт представлен плотно прилегающими друг к другу клетками, мигрирующими пластом в меридиональном направлении по внутренней поверхности перидермы. Перемещение эпибласта происходит с некоторой задержкой по сравнению с гипобластом. Мигрирующие к вегетативному полюсу пласты оставляют на поверхности желтка клетки, которые образуют покров желточного мешка и в скором будущем его сосудистую систему.  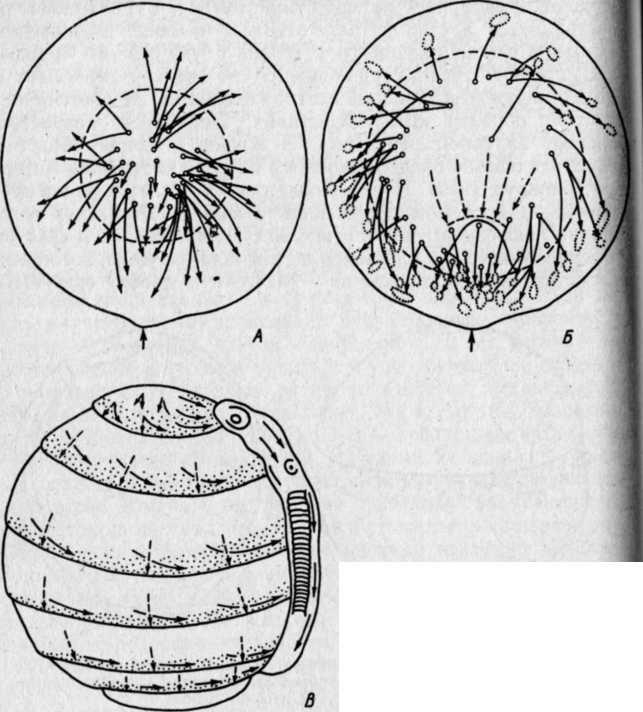 Рис. VIII.4. Морфогенетические движения клеток у лосося (по Ballard 1973). А – перемещение глубоких клеток из центра бластодермы; Б – образование краевого узелка; В – пути миграций клеток эпибласта (пунктирные стрелки) и гипобласта (сплошные стрелки) Презумптивные зачатки. На протяжении гаструляции глубокие клетки бластодермы движутся по своим траекториям к зародышевому щитку. Достигнув его, клетки рассортировываются и занимают определенное положение: верхнюю часть зародыша представляет эктодерма, среднюю – мезодерма и материал хорды, нижний пласт, лежащий на перибласте, представлен энтодермой (рис.VIII.5). Это свидетельствует о том, что в раннем эмбриогенезе, на бластуле, происходит детерминация этих клеток. Она возникает, очевидно, в результате качественных различий клеток, расположенных в разных участках бластодермы.   Рис. VIII.5. Срез через зародышевый щиток карпа (ориг.). 1 – перидерма, 2 – эктодерма, 3 – мезодерма, 4 – энтодерма, 5 – перибласт, 6 – желток Современные карты презумптивных зачатков составлены для стальноголового лосося S. gairdneri и представителя сем. чукучановых – катостомуса Catostomus commersoni Баллардом (Ballard, 1981, 1982) на основании экспериментальных данных.  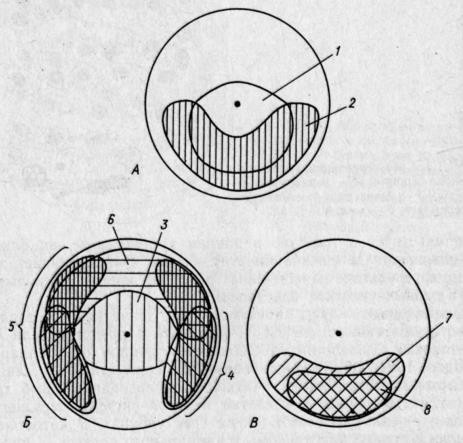 Рис. VIII.6. Карта презумптивных зачатков катостомуса (по Ballard, 1982). А – верхний слой бластодермы: 1 – передний и средний мозг, 2 – задний мозг. Б – средний слой бластодермы: 3 – головная мезодерма, 4 – передние туловищные сомиты, 5 – туловищные сомиты после 10-го сомита, 6 – хвостовые сомиты. В – нижний слой бластодермы: 7 – энтодерма, 8 – хорда Карты презумптивных зачатков у исследованных рыб трехмерны. Это означает, что клетки нервной системы, мезодермы, Хорды и кишки располагаются на трех уровнях. У катостомуса при рассмотрении бластодермы с анимального полюса (в нижней части бластодермы находится место закладки зародышевого узелка) почти весь ее верхний слой занимает презумптивная центральная нервная система и покровы, причем среднюю часть – передний и средний мозг, а изогнутую в виде широкого полумесяца нижнюю часть – задний мозг (рис.VIII.6, А). В самом глубоком слое нижней части бластодермы находится презумптивная энтодерма, а над ней, занимая несколько меньшую область материал хорды (рис.VIII.6, В). Всю заднюю толщу бластодермы и центральную часть между презумптивными нервной системой и хордой занимает мезодерма. Причем, в центральной части располагается головная мезодерма, позади нее материал хвостовых сомитов. С боков передней части бластодермы, в виде овалов – передние туловищные сомиты, с боков же в виде широких длинных полулуний – туловищные сомиты после 10-го сомита (рис.VIII.6, Б). В бластодерме лосося расположение презумптивных зачатков в общих чертах сходно с таковым у катостомуса. Приводившиеся ранее в литературе карты презумптивны зачатков были составлены Пастельсом (Pasteels, 1936) для лосося и Оппенгеймер (Oppengeimer, 1936) для фундулуса. Эти карты были одномерны. По существовавшим до работ Балларда представлениям, гаструляция у костистых рыб происходит по общему для позвоночных животных плану, т. е. благодаря инвагинации клеточного материала, из которого впоследствии дифференцируются зародышевые листки. этап. ОрганогенезК началу органогенеза тело зародыша в виде валика располагается на желточном мешке (рис.VIII.7, А). Высота тела, особенно передней части, на протяжении этапа заметно возрастает. Дифференциация органов происходит в краниально-каудальном направлении. Завершается этап образованием зачатка хвостового отдела – хвостовой почки. Дифференциация хорды в осевом зачатке мезодермы происходит на ранней стадии развития, еще до сегментации туловищной мезодермы. Клетки хорды отделяются от мезодермальных в виде тяжа по оси зародыша. Структура клеток хорды и мезодермы вначале сходна. Но вскоре клетки хорды уплощаются, вакуолизируются и приобретают форму дисков (Laale, 1982). Дифференциация мезодермы. В самом начале этапа под головным отделом зародышей в виде валиков располагается головная мезодерма и мезодерма передних туловищных сомитов (рис.VIII.7, А). Головная мезодерма далее утончается и распадается на мезенхимные клетки. Мезодерма передних сомитов в виде двух тяжей продолжает продвигаться в каудальном направлении. Туловищная мезодерма, мигрировавшая из зародышевого кольца справа и слева от оси зародыша, образует два пласта, сливающихся с мезодермой, поступившей в зародыш спереди. Мезодерма располагается в виде пластов по обе стороны от хорды, представлена утолщенной верхней частью и суженной средней частью, которая переходит в тонкий вентральный слой – спланхнотом, или боковуюпластинку. Сегментации, т. е. расчленению на отдельные блоки – сомиты, подвергается лишь утолщенная дорсальная часть мезодермы. При этом происходит объединение мезодермальных клеток в группы, разделенные узкими щелями. Число сомитов постепенно возрастает, благодаря обособлению их из задней несегментированной части мезодермы (рис.VIII.7, Б–Г).  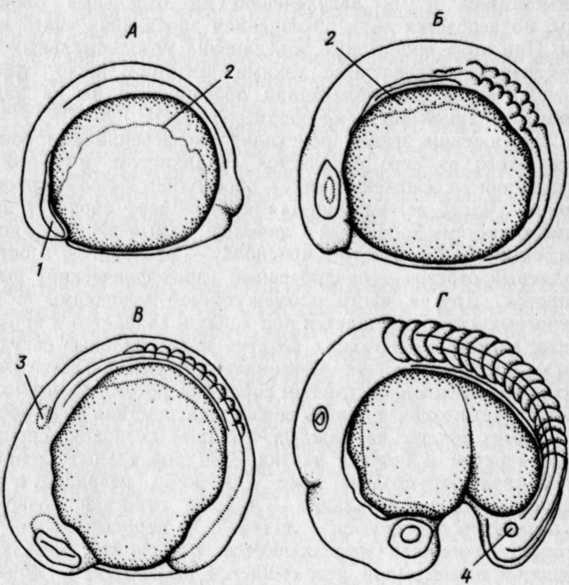 Рис. VIII.7. V этап – органогенез у карпа (ориг.). А–закладка зародыша (около 10 ч), 5 – появление зачатков глаз (13 ч), В – появление слуховых плакод (15 ч), Г –появление в глазах хрусталиков, полостей в слуховых зачатках и купферова пузырька (18–20 ч). 1 – головная мезодерма, 2 – боковая пластинка, 3 – слуховая плакода, 4 – купферов пузырек На протяжении этапа происходит дифференциация сомита. Прежде всего от него отделяется спланхнотом, и в области его отделения от «ножек сомита» обособляется промежуточная мезодерма. Часть ее, называемая нефротомом, образует выделительную систему зародыша и личинки. Самый передний короткий отрезок тяжа формирует предпочку – pronephros, а остальной длинный отрезок – предпочечный (пронефрический) выводной проток. Другая часть промежуточной мезодермы в виде мезенхимных клеток смещается под хорду и сливается в продольный тяж клеток, образующих позднее магистральные сосуды спинную аорту и задние кардинальные вены. Третья часть мезенхимных клеток представляет собой склеротомную мезенхиму. Она мигрирует к хорде и охрящевевает впоследствии на ее оболочке в четырех местах на каждый будущий позвонок: справа и слева в верхней и нижней частях. Два дорсальных хрящевых зачатка образуют позднее, уже у личинок, невральную дугу, позвонка, а два вентральных – нижние отростки позвонков. Тела позвонков образуются в личиночном периоде жизни. Еще некоторое количество отделяющейся от сомита мезенхимы, называемой дерматомной, перемещается под эпителий и образует соединительнотканный слой кожи. После отделения от сомитов нефро-, склеро- и дерматома, а также мезенхимы кровеносных сосудов остается их основная часть – миотомы, которые образуют туловищную мускулатуру (Иванов, 1945). Сбоку сомиты и образующиеся после их дифференциации миотомы имеют вначале форму прямоугольников со сглаженными углами. При образовании примерно половины туловищных миотомов форма их меняется. Они изгибаются в средней части и приобретают вид конусов, направленных вершиной вперед (рис.VIII.7, Г). В связи с развитием миофибрилл миотомы начинают сокращаться, что приводит к легким изгибам тела зародышей. Дифференциация энтодермы. Под мезодермальным слоем зародыша располагается тонкий слой энтодермы, лежащей на перибласте. Задняя часть энтодермального зачатка расширена и в ней примерно в середине этапа появляется характерный для костистых рыб провизорный орган – купферов пузырек (рис.VIII.7, Г), названный в честь описавшего его немецкого эмбриолога Купфера. Этот пузырек представляет собой маленькое расширение в энтодерме. С удлинением энтодермального зачатка он перемешается в каудальном направлении, затем уменьшается и исчезает. Функция купферова пузырька до сих пор не ясна. Наблюдения показывают, что возникает он в заднем отделе зачатка кишки и перемещается вместе с ним. В месте исчезновения пузырька образуется анальное отверстие (Крыжановский, 1956). Дифференциация эктодермы. Верхний эктодермальный слой зародыша имеет утолщенную посередине невральную пластинку. Клетки ее, конвергируя к центру, образуют плотный нервный тяж, утолщенный в головном отделе. По мере роста зародыша происходит удлинение неврального тяжа. Зачаток головного мозга, представленный на ранних стадиях одним утолщением, с началом сегментации мезодермы дифференцируется на три отдела. В них вскоре появляются полости, и плотные зачатки превращаются в мозговые пузыри – передний, средний и задний – соответственно – prosencephalon, mesencephalon, rhombencephalon. Зачатки глаз у рыб овальной формы, плотные, они появляются у зародышей в области переднего мозгового утолщения на начальных стадиях сегментации мезодермы (у карповых рыб при 4–5 сегментах), когда полости в зачатке мозга еще не образовались (рис. VIII. 7, Б). Вскоре в зачатках глаз появляются щелевидные полости, и они превращаются в глазные пузыри. На более поздних стадиях развития полости в глазах увеличиваются. Началом превращения глазных пузырей в бокалы считают утолщение наружных стенок глазных пузырей и их инвагинацию вовнутрь. Одновременно закладываются хрусталики в виде утолщения внутреннего слоя эпителия, прилегающего с боковых сторон к глазу. К концу этапа хрусталики отделяются от эпителия и размещаются в полости глаза (рис. VIII. 7, Г). Глаза остаются связанными с мозгом широкими глазными стебельками (Бабурина, 1972). Слуховые зачатки в виде плакод появляются у зародышей на стадии 8–12 туловищных сегментов в области заднего мозгового пузыря (рис. VIII. 7, В). Вскоре в них благодаря расхождению меток возникают узкие, постепенно увеличивающиеся полости и плакоды превращаются в слуховыепузырьки(рис. VIII. 7, Г), Стенки их с увеличением полости утончаются, но в местах расположения будущих чувствующих пятен остаются утолщенным (Макеева, 1958). Расстояние между слуховыми пузырьками и глазами на этом этапе довольно большое. Оно равно ширину 7–8 туловищных сегментов. Это связано с тем, что головной мозг не имеет изгибов и лежит в одной плоскости. За слуховыми плакодами у рыб закладываются, также в виде плакод, зачатки органов боковой линии. Они разрастаются под эпидермисом вперед – над и под глазами, и назад, в туловище. По мере разрастания зачаток оставляет местные утолщения, из которых впоследствии развиваются чувствующие бугорки – невромасты (Дислер, 1960; Смирнов, 1985). |