Стероидные гормоны. Стероидные гормоны
 Скачать 220.5 Kb. Скачать 220.5 Kb.
|

Биологическое действие стероидных гормоновСтероидные гормоны яичников (эстрогены, прогестерон, андрогены, кортикостероиды) влияют на обмен веществ не только в половых органах, но и в других системах. В нервной системе гормоны влияют непосредственно на нервные клетки. Под воздействием гормонов изменяется течение рефлекторных реакций, возникающих в различных системах организма. Эти гормоны принимают активное участие в осуществлении адаптационных реакций организма при различных физиологических процессах и вредных воздействиях. Механизм действия гормонов в значительной степени определяется и функциональным состоянием рецепторов в различные физиологические периоды жизни женщины, и функцией высших нервных центров. Действие гормонов опосредуется через нервную систему; особое значение имеет симпатический отдел вегетативной нервной системы. Симпатические нервные волокна имеют в гладких мышечных волокнах матки а- и р-рецепторы. а-Рецепторы воспринимают симпатические сосудосуживающие импульсы, а также импульсы, сокращающие мышцы; они более чувствительны к эстрогенам. р-Рецепторы воспринимают сосудорасширяющие импульсы, а также импульсы, вызывающие расслабление мышц. р-Рецепторы более чувствительны к прогестерону. В связи с этим активность а- и р-рецепторов меняется в различных фазах менструального цикла, в процессе беременности и родов. Так, в фазе пролиферации эндометрия при относительном преобладании эстрогенов преобладает чувствительность ос-рецепторов; в фазе секреции при преобладании прогестерона доминирует активность р-рецепторов; то же происходит во время беременности, когда подавляются сокращения матки. Во время родов активизируются а-рецепторы, что способствует сокращению матки. В последние годы предложены вещества, избирательно блокирующие а- или р-рецепторы мышечных волокон матки — так называемые адреноблокаторы (рис. 10). Это открывает большие перспективы в борьбе с акушерской патологией, обусловленной аномалиями родовой деятельности. Половые гормоны яичников не действуют изолированно, а вместе с другими гормонами (гипофиза, надпочечников и щитовидной железы) участвуют в регуляции жизненных функций. При изменении секреции одного гормона возникают цепные нейрогормональные реакции, приводящие к изменению гормональной активности других эндокринных желез. Особенно это относится к кортикостероидной функции надпочечников и щитовидной железе. Эстрогены. Специфическими особенностями эстрогенных влияний в организме женщины являются стимуляция пролифе-ративных процессов, улучшение кровоснабжения органов-эффекторов, стимуляция развития вторичных половых признаков. 1. Влияние на половые органы. Особенно отчетливо проявляется влияние эстрогенных гормонов на матку. Они оказывают прямое действие на мышечной слой, слизистую оболочку и сосуды матки, улучшают ее кровоснабжение, вызывают пролиферацию эндо- и миометрия, в период полового созревания способствуют увеличению размеров матки, ее росту до величины, соответствующей размеру детородного возраста. Прекращение эстрогенных влияний в старости приводит к уменьшению размера матки, истончению мышечного слоя и эндометрия. Циклические колебания уровня эстрогенных гормонов в течение менструального цикла у женщин после периода полового созревания вызывают закономерные изменения в строении эндометрия— фазу пролиферации. Происходят пролиферация же-3 эндометрия, рост и уплотнение его стромы. Железы эндо метрия приобретают удлиненную форму, увеличивается количество митозов, в клетках желез появляется щелочная фосфатаза, увеличивается содержание рибонуклеопротеидов, усиливается кровоснабжение эндометрия. Стимулируются рост и секреторная активность шеечного эпителия. Слизистая оболочка матки подготовляется к переходу в секреторную фазу. В клиническом эксперименте вызвать секреторные изменения в эндометрии с помощью гестагенов без предварительного эст-рогенного воздействия не удается. Эстрогены способствуют регенерации эндометрия после отторжения его во время менструации. В мышце матки повышается чувствительность к воздействию окситоцических веществ, отмечается повышенное выделение ацетилхолина. В маточных трубах происходят аналогичные, но менее выраженные изменения: улучшается кровообращение, наблюдается пролиферация слизистой оболочки, усиливается мышечная активность. Во влагалище также улучшается кровоснабжение, стимулируется процесс ороговения многослойного плоского эпителия, а в слизистой оболочке откладывается гликоген. В яичнике эстрогены стимулируют развитие гранулезных клеток. 2. Влияние на молочные железы. Эстрогенные гормоны способствуют росту молочных желез (в период полового созревания повышают их кровоснабжение), росту секреторного эпителия, преимущественно в молочных ходах. 3. Влияние на гипоталамо-гипофизарный отдел центральной нервной системы. Эстрогены в избыточном количестве тормозят фолликулярную активность яичника и непосредственно, и через гипоталамус—-гипофиз. Очень большие дозы эстрогенов вызывают атрофию яичников, небольшие количества их стимулируют гормонообразование (ФСГ) в гипофизе, большие — тормозят его. Эстрогены блокируют образование СТГ. Малые дозы эстрогенов, по мнению большинства авторов, стимулируют гормонообразование в щитовидной железе, большие — блокируют его. Эстрогены способствуют атрофии эпифиза. Уровень содержания эстрогенов в крови обеспечивает так называемый стероидный контроль высших регулирующих органов. 4. Общее влияние эстрогенов на организм. Эстрогены оказывают влияние на нервную и сердечно-сосудистую системы, а также на обмен веществ. Они способствуют задержке в организме азота, натрия, жидкости, в костях — кальция, фосфора и тем самым образованию костного вещества. В то же время в старческом возрасте снижение эстрогенного уровня может иногда способствовать развитию остеопороза. Под воздействием эстрогенов в эпителии мочевыделительной системы (уретра, мочевой пузырь, мочеточники) возникают специфические циклические изменения. В связи с этим динамическое исследование осадка мочи во время менструального цикла позволяет судить о функции яичников (см. главу III). К факторам, регулирующим действие эстрогенов, относятся фолиевая кислота и реактивность тканей-мишеней. Гестагены. Гестагены, оказывая влияние на половые органы, стимулируют секреторное превращение эндометрия, в котором имеются пролиферативные изменения, и децидуальную реакцию стромы. После прекращения влияния гестагенов наступает Дёсквамация эндометрия и начинается кровотечение. Гестагены, так же как и эстрогены, спососбтвуют гипертрофии мышечного слоя матки и увеличению амплитуды отдельных сокращений; число спонтанных маточных сокращений снижается, одновременно повышается тонус мускулатуры перешейка (при беременности — нижнего маточного сегмента) и шейки матки. Механизм тормозящего влияния гестагенов на миомет-рий не совсем ясен. Высказывается предположение, что это является результатом блокирования передачи нервных импульсов с одного мышечного волокна на другое вследствие изменения содержания в клетках электролитов; под влиянием прогестерона снижается внутриклеточная концентрация ионов калия, необходимая для сокращения мышцы матки [Czapo, 1963]. v-лизь в шеечном канале теряет способность кристаллизоваться в виде «папоротника» под влиянием прогестерона. 3-255 зз 1. Под влиянием прогестерона поверхностные клетки влагалищного эпителия слущиваются и проявляется большое количество клеток с завернутыми краями и зернистой протоплазмой. Количество ороговеваюших клеток снижается до 18—20, и преобладают промежуточные клетки. ‘ Сокращение маточных труб, так же как и мышцы матки, возникают реже, приобретают более интенсивный характер. В яичниках гестагены удлиняют выделение эстрогенов и тормозят овуляцию, возможно, путем подавления ЛГ. Вследствие специфического влияния на половые органы, главным образом на матку, гестагены называют «протекторами» беременности, так как они’подготовляют эндометрий к имплантации оплодотворенного плодного яйца и делают матку нечувствительной к действию окситоцических веществ.
- 4. Общее влияние гестагенов на организм заключается в выраженном воздействии на обменные процессы. Оказывая диуретическое влияние, они способствуют выделению натрия и жидкости из организма без выведения калия. Наиболее характерной особенностью действия прогестерона является его термогенный эффект, заключающийся в том, что при увеличении уровня прогестерона в организме повышается температура тела.* На этом основан тест двухфазного характера базальной температуры в течение менструального цикла, что принято объяснять прямым действием на гипоталамус.’Термо- генный эффект прогестерона доказывается также повышением температуры тела на 0,5—1 °С у кастрированных женщин после дачи им 80 мг прегненолона ежедневно [Israel, 1967]. __ Андрогены. Андрогены у женщин секретируются корой над- почечников и яичниками. В яичниках андрогены образуются в процессе стероидогенеза в незначительном количестве; глав- ным источником образования андрогенов является сетчатая зона коры надпочечника. Данные последних лет о повышении экскреции тестостерона во второй фазе менструального цикла позволяют предположить, что тестостерон секретируется также желтым телом. Андрогены влияют на активность различных ферментов. Так, введение андрогенов повышает активность дегидрогеназы, янтарной кислоты и цитохромоксидазы в семенных пузырьках и предстательной железе кастрированных животных. Ренотроп-ная активность андрогенов также связана с увеличением активности аргиназы и уменьшением активности щелочной фосфата-зы в почках. Андрогены способствуют задержке азота в организме, стимулируя синтез белка и снижая скорость катаболизма аминокислот, активно понижают выведение мочевины. Андрогены ускоряют рост костей и окостенение эпифизарных хрящей. Биологическая роль андрогенов начинает проявляться тогда, когда они начинают вырабатываться в яичниках или в коре надпочечников. Печень не только связывает тестостерон с последующей экскрецией его почками и кишечником, но и переводит андростендион и дегидроэпиандростерон в тестостерон. Известно, что андрогены обладают анаболическим свойством. Это демтш1е_о_б^^лов^п^ается повышением синтеза.-белка на рибосомальноп РНК, что приводит к задержке азота. Андрогены вызывают накопление фосфора и калия, задержку натрия и хлора, понижают выделение мочевины. При избыточном выделении андрогенов — гормонов с диаметрально противоположным действием, чем эстрогены, вначале проявляются признаки дефеминизации, а затем маскулинизации. Для стероидных гормонов и витамина D в цитоплазме соответствующих клеток-мишеней имеются специфические рецепторы. Наличие рецепторов в других органеллах, таких как митохондрии или мембрана ядра, для этих гормонов до сих пор не доказано. В ядре находятся специфические рецепторы для стероидных гормонов, витамина D и гормонов щитовидной железы.
|
 |
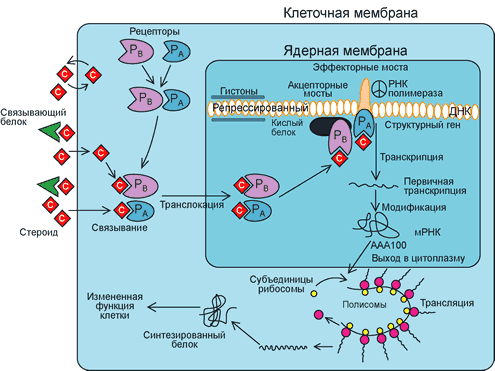
Хотя для всех 5 физиологически определяемых классов стероидных гормонов (эстрогены, прогестерон, андрогены, глюкокортикоиды, минералокортикоиды) найдены специфические рецепторы, каждый стероидный гормон может реагировать с рецепторами других классов стероидов. Так, андрогены при высоких концентрациях могут конкурировать с эстрогенами за связывание достаточно высокоспецифичными рецепторами эстрогенов. Рецепторы андрогенов вообще проявляют высокое сродство к большому числу стероидов. Прогестагены могут имитировать эффекты андрогенов, потенцируя или ингибируя их действие. Например, прогестерон является слабым андрогеном при действии на клетки предстательной железы, в то время как производное прогестерона, ципротеронацетат, действует в качестве антиандрогена. На клетки гепатомы прогестерон действует как антиглюкокортикоид, а на почки — как минералокортикоид. Большей или меньшей специфичностью к рецепторам обладают также аналоги стероидов и синтетические вещества нестероидной природы, но обладающие активностью стероидов.
При действии стероидных гормонов на клетки-мишени происходит отчетливое изменение числа рецепторов. Например, при действии физиологических доз эстрадиола число цитозольных рецепторов, которое составляет примерно 20000—40000 на клетку матки, снижается приблизительно наполовину. Все ли рецепторные комплексы проникают в ядро клетки, не ясно, но многие тысячи таких комплексов могут там находится. Уменьшение содержания цитозольных рецепторов стимулирует их новый синтез, в результате чего через 18 — 24 ч после этого достигается исходный уровень. Стероиды регулируют не только концентрацию своих собственных рецепторов, но и модулируют также концентрацию рецепторов других стероидов. Так, эстрогены стимулируют рецепторы для прогестерона, в то время как прогестерон, как правило, понижает концентрацию рецепторов эстрогенов в репродуктивном тракте. Рецепторы андрогенов регулируются эстрогенами: например, в яйцеводах кур эстрогены повышают содержание рецепторов андрогенов примерно в 10 раз.
Поскольку связывание цитозольных рецепторов со стероидами (оккупация рецептора) является предпосылкой для функции гормонозависимых клеток, то можно сделать выводы, имеющие определенное практическое значение для терапии гормонозависимых опухолей. Это относится, например, к раку молочной железы. Примерно 25 — 30% больных с прогрессирующим раком молочной железы обнаруживают объективную ремиссию при эндокринной терапии. Известно, что примерно у 90% больных, у которых содержание рецепторов эстрогенов в опухолях молочной железы выше определенной величины (около 30% всех опухолей молочной железы), эндокринная терапия (овариэктомия и адреналэктомия, гипофизэктомия или применение больших доз андрогенов, эстрогенов или антиэстрогенов) может давать ремиссию. Опухоли с большим количеством рецепторов имеют меньшую вероятность и быстроту рецидива, чем те, у которых мало рецепторов.
Наряду с упомянутым механизмом действия стероидов новые работы на ооцитах амфибий показывают, что, вероятно, стероидные гормоны могут проявлять свое действие через поверхность клетки, а следовательно, без проникновения в ее ядро. Процесс, видимо, протекает с ингибированием мембранно-связанной аденилатциклазы, причем предполагают прямое и непрямое действие на нуклеотидрегулирующую субъединицу.