тканевое дых. Тканевое дыхание
 Скачать 337.19 Kb. Скачать 337.19 Kb.
|

|
Тканевое дыхание, клеточное дыхание, совокупность ферментативных процессов, протекающих при участии кислорода воздуха в клетках органов и тканей, в результате чего продукты расщепления углеводов, жиров, белков окисляются до углекислого газа и воды, а значит, часть освобождающейся энергии запасается в форме богатых энергией, или макроэргических соединений. Т. д. отличают от внешнего дыхания — совокупности физиологических процессов, обеспечивающих поступление в организм кислорода и выведение из него углекислого газа. Многие ферменты, катализирующие эти реакции, находятся в особых клеточных органоидах — митохондриях. На все проявления жизни — рост, движение, раздражимость, самовоспроизведение и др. — организм расходует энергию. Формой энергии, пригодной для использования клетками, является энергия химических связей (главным образом фосфатных) в макроэргических соединениях — аденозинтрифосфорной кислоте (АТФ) и др. Для синтеза АТФ необходим приток энергии извне. По способам извлечения энергии существует принципиальное различие между автотрофными организмами и гетеротрофными организмами. Клетки зелёных растений — наиболее типичных автотрофов — в процессе фотосинтеза используют энергию солнечного света для синтеза АТФ и глюкозы. (Образование из глюкозы более сложных молекул происходит в клетках растений также в процессе Т. д.) В клетках гетеротрофов — животных и человека — единственным источником энергии является энергия химических связей молекул пищевых веществ. Молекулы различных соединений, выполняющие роль биологического «топлива» (глюкоза, жирные кислоты, некоторые аминокислоты), образовавшись в клетках животного организма или поступив в кровь из пищеварительного тракта, претерпевают ряд последовательных химических превращений. В процессе Т. д. можно наметить три основные стадии: 1) окислительное образование ацетилкофермента А (активная форма уксусной кислоты) из пировиноградной кислоты (промежуточный продукт расщепления глюкозы), жирных кислот и аминокислот; 2) разрушение ацетильных остатков в трикарбоновых кислот цикле с освобождением 2 молекул углекислого газа и 4 пар атомов водорода, частично акцептируемых коферментами никотинамидадениндинуклеотидом и флавинадениндинуклеотидом и частично переходящих в раствор в виде протонов; 3) перенос электронов и протонов к молекулярному кислороду (образование H2O) — процесс, катализируемый набором дыхательных ферментов и сопряжённый с образованием АТФ (так называемое окислительное фосфорилирование). Первые две стадии подготавливают третью, в ходе которой в результате последовательных окислительно-восстановительных реакций происходит освобождение основной части энергии, вырабатываемой в клетке. При этом около 50% энергии в результате окислительного фосфорилирования запасается в форме богатых энергией связей АТФ, а остальная часть её выделяется в виде тепла. Т. д. обеспечивает образование и постоянное пополнение АТФ в клетках. В случае недостатка в снабжении клеток животных и человека кислородом запасы АТФ не исчерпываются сразу. Их пополнение может происходить в результате включения дополнительных механизмов — систем анаэробного (без участия кислорода) распада углеводов — гликолиза и гликогенолиза. Однако этот путь энергетически во много раз менее эффективен и не может обеспечить функции и целостность структуры органов и тканей. Биологическая роль Т. д. не исчерпывается существенным вкладом в энергетический обмен организма. На различных его этапах образуются молекулы органических соединений, используемых клетками в качестве промежуточных продуктов для различных биосинтезов ДЫХАНИЕ ТКАНЕВОЕ (син. клеточное дыхание) - совокупность окислительно-восстановительных процессов в клетках, органах и тканях, протекающих с участием молекулярного кислорода и сопровождающихся запасанием энергии в молекулах АТФ. В процессе Т.д. можно наметить три основные стадии: 1) окислительное образование ацетилКоФ-А (активная форма уксусной кислоты) из пировиноградной кислоты (промежуточный продукт расщепления глюкозы), жирных кислот и аминокислот; 2) разрушение ацетильных остатков в ЦТК с освобождением 2 молекул углекислого газа и 4 пар атомов водорода, частично акцептируемых КоФ – НАД и ФАД и частично переходящих в раствор в виде протонов; 3) перенос электронов и протонов к молекулярному кислороду (образование H2O) — процесс, катализируемый набором дыхательных ферментов и сопряжённый с образованием АТФ (так называемое окислительное фосфорилирование). Первые две стадии подготавливают третью, в ходе которой в результате последовательных окислительно-восстановительных реакций происходит освобождение основной части энергии, вырабатываемой в клетке. При этом около 50% энергии в результате окислительного фосфорилирования запасается в форме богатых энергией связей АТФ, а остальная часть её выделяется в виде тепла. 3) При переносе электронов в митохондриальной мембране каждый комплекс дыхательной цепи направляет свободную энергию окисления на перемещение протонов (положительных зарядов) через мембрану, из матрикса в межмембранное пространство, что приводит к образованию разности потенциалов на мембране: положительные заряды преобладают в межмембранном пространстве, а отрицательные - со стороны матрикса митохондрий. При достижении определенной разности потенциалов (220 мВ) белковый комплекс АТФ-синтетазы начинает транспортировать протоны обратно в матрикс, при этом превращает одну форму энергии в другую: образует АТФ из АДФ и неорганического фосфата. Так происходит сопряжение окислительных процессов с синтетическим - с фосфорилированием АДФ. Пока происходит окисление субстратов, пока происходит перекачка протонов через внутреннюю митохондриальную мембрану - идет сопряженный с этим синтез АТФ, то есть окислительное фосфорилирование. Энергия, выделяющаяся в ходе этих реакций, трансформируется в трансмембранный протонный потенциал. Фермент АТФ-синтаза использует этот градиент для синтеза АТФ, преобразуя его энергию в энергию химических связей. Подводя итоги энергетики процесса дыхания, подсчитаем, сколько всего молекул АТФ может образоваться при распаде одной молекулы глюкозы. В первую анаэробную фазу дыхания при распаде одной молекулы глюкозы до двух молекул пировиноградной кислоты в процессе субстратного фосфорилирования накапливаются две молекулы АТФ. Одновременно на этой фазе дыхания при окислении ФГА до ФГК в цитозоле образуются две молекулы восстановленных коферментов (2 НАД-Н2). Они диффундируют через наружную мембрану и окисляются в дыхательной цепи благодаря наличию у растений НАД-Н-дегидрогеназы, локализованной на наружной поверхности внутренней мембраны. При этом синтезируется 6 молекул АТФ. В аэробной фазе дыхания при окислении пировиноградной кислоты образуются 4НАД-Н2. Их окисление в дыхательной цепи приводит к образованию 12 АТФ. Кроме того, в цикле Кребса восстанавливается одна молекула флавиновой дегидрогеназы (ФАД-Н2). Окисление этого соединения в дыхательной цепи приводит к образованию 2 АТФ, поскольку одно фосфорилирование не происходит. При окислении молекулы а-кетоглутаровой кислоты до янтарной энергия непосредственно накапливается в одной молекуле АТФ (субстратное фосфорилирование). Таким образом, окисление одной молекулы пировиноградной кислоты сопровождается образованием ЗСО2 15 молекул АТФ. Однако при распаде молекулы глюкозы образовались две молекулы пировиноградной кислоты. Следовательно, всего в аэробной фазе дыхания образуется 6 молекул СО2 и 30 молекул АТФ плюс 8АТФ в анаэробной фазе. Итого 6 молекул СО2 и 38 молекул АТФ образуется в процессе окислительно-дыхательного распада молекулы гексозы. Дыхательная цепь Дыхательная цепь является частью процесса окислительного фосфорилирования ). Компоненты дыхательной цепи катализируют перенос электронов от НАДН + Н+ или восстановленного убихинона (QH2) на молекулярный кислород. Из-за большой разности окислительно-восстановительных потенциалов донора (ия является высокоэкзергонической Большая часть выделяющейся при этом энергии используется для создания градиента протонов (см. с. 1НАДН + Н+ и, соответственно, QH2) и акцептора (О2) реакц и, наконец, для образования АТФ с помощью АТФ-синтазы. А. Компоненты дыхательной цепи Дыхательная цепь включает три белковых комплекса (комплексы I, III и IV), встроенных во внутреннюю митохондриальную мембрану, и две подвижные молекулы-переносчики — убихинон (кофермент Q) и цитохром с. Сукцинатдегидрогеназа, принадлежащая собственно к цитратному циклу, также может рассматриваться как комплекс II дыхательной цепи. АТФ-синтаза иногда называется комплексом V, хотя она не принимает участия в переносе электронов. Комплексы дыхательной цепи построены из множества полипептидов и содержат ряд различныхокислительно-восстановительных коферментов, связанных с белками . К ним принадлежат флавин [ФМН (FMN) или ФАД (FAD), в комплексах I и II], железо-серные центры (в I, II и III) и группы гема (в II, III и IV). Детальная структура большинства комплексов еще не установлена. Электроны поступают в дыхательную цепь различными путями. При окислении НАДН + Н+ комплекс I переносит электроны через ФМН и Fe/S-центры на убихинон. Образующиеся при окислении сукцината, ацил-КоА и других субстратов электроны переносятся на убихинон комплексом II или другой митохондриальной дегидрогеназой через связанный с ферментом ФАДН2 или флавопротеин , При этом окисленная форма кофермента Q восстанавливается в ароматическийубигидрохинон. Последний переносит электроны в комплекс III, который поставляет их через два гема b, один Fe/S-центр и гем с1 на небольшой гемсодержащий белок цитохром с. Последний переносит электроны к комплексу IV, цитохром с-оксидазе. Цитохром с-оксидаза содержит для осуществления окислительно-восстановительных реакций два медьсодержащих центра (CuA и CuB) и гемы а и а3, через которые электроны, наконец, поступают к кислороду. При восстановлении О2 образуется сильный основной анион О2-, который связывает два протона и переходит а воду. Поток электронов сопряжен с образованным комплексами I, III и IV протонным градиентом. Б. Организация дыхательной цепи Перенос протонов комплексами I, III и IV протекает векторно из матрикса в межмембранное пространство. При переносе электронов в дыхательной цепи повышается концентрация ионов H+, т. е. понижается значение рН. В интактных митохондриях по существу только АТФ-синтаза позволяет осуществить обратное движение протонов в матрикс. На этом основано важное в регуляторном отношении сопряжение электронного переноса с образованием АТФ Как уже упоминалось, все комплексы с I по V интегрированы во внутренней мембране митохондрий, тем не менее обычно они не контактируют друг с другом, так как электроны переносятся убихиноном и цитохромом с. Убихинон благодаря неполярной боковой цепи свободно перемещается в мембране. Водорастворимый цитохром с находится на внешней стороне внутренней мембраны. Окисление НАДН (NADH) комплексом I происходит на внутренней стороне мембраны, а также в матриксе, где происходит также цитратный цикл и β-окисление — самые важные источники НАДН. В матриксе протекают, кроме того, восстановление O2 и образование АТФ (ATP). Полученный АТФ переносится по механизму антипорта (против АДФ) в межмембранное пространство , откуда через порины проникает в цитоплазму. Перенос электронов от НАДН к молекулярному кислороду через электронно-транспортную цепь (ЭТЦ), обеспечивающий процесс фосфорилирования АДФ с образованием АТФ, получил название окислительного фосфорилирования. На этой стадии клеточного дыханияприсутствие кислорода является обязательным условием. Окислительное фосфорилирование протекает на внутренней мембране митохондрий, в которой находятся белки-переносчики электрон-транспортной цепи переноса электронов (дыхательной цепи) и комплексы АТФ-синтетазы Элементы дыхательной цепи последовательно окисляются и восстанавливаются, в результате чего энергия высвобождается небольшими порциями и используется для перекачивания протонов (H+) из матрикса митохондрии через внутреннюю митохондриальную мембрану в межмембранное пространство и далее за пределы митохондрии (при этом они используются для синтеза АТФ). Таким образом, дыхательная цепь, или цепь переноса электронов в митохондрии, является главной системой превращения энергии в химические связи АТФ. При полном окислении одной молекулы глюкозы обеспечивается в конце концов образование примерно 36 высокоэнергетических фосфатных связей в виде АТФ. Большая часть АТФ (30 молекул) образуется в процессе окислительного фосфорилирования.    ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ, синтез АТФ из аденозиндифосфата и неорг. фосфата, осуществляющийся в живых клетках, благодаря энергии, выделяющейся при окислении орг. в-в в процессе клеточного дыхания. В общем виде окислительное фосфорилирование и его место в обмене в-в можно представить схемой: 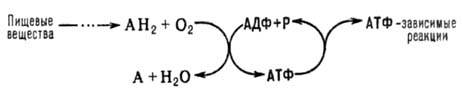 АН2-орг. в-ва, окисляемые в дыхат. цепи (т. наз. субстраты окисления, или дыхания), АДФ-аденозиндифосфат, Р-неорг. фосфат. Поскольку АТФ необходим для осуществления мн. процессов, требующих затраты энергии (биосинтез, совершение мех. работы, транспорт в-в и др.), Окислительное фосфорилирование играет важнейшую роль в жизнедеятельности аэробных организмов. Образование АТФ в клетке происходит также благодаря др. процессам, напр. в ходе гликолиза и разл. типов брожения, протекающих без участия кислорода. Их вклад в синтез АТФ в условиях аэробного дыхания составляет незначит. часть от вклада окислительного фосфорилирования (ок. 5%). |