Транспорт воды через аквопорины мембраны клетки. Транспорт воды через аквапорины мембраны клетки. Транспорт воды через аквапорины мембраны клетки
 Скачать 218.5 Kb. Скачать 218.5 Kb.
|

|
Транспорт воды через аквапорины мембраны клетки https://meduniver.com/Medical/genetika/transport_vodi_cherez_akvaporini.html Аквапорины обеспечивают быстрый и селективный транспорт молекул воды через мембраны клетки Аквапорины являются тетрамерами, состоящими из четырех идентичных субъединиц, каждая из которых образует пору. Селективный фильтр аквапорина характеризуется тремя главными особенностями (фильтрация по размеру, электростатическое отталкивание и ориентация диполей воды), которые обеспечивают его высокую селективность по отношению к воде. Для протекания многих физиологических процессов необходим транспорт воды через клеточные мембраны. Однако пассивная диффузия воды через липидный бислой биологических мембран не поддается регуляции и характеризуется ограниченной проницаемостью и низкой скоростью. Быстрый и селективный транспорт воды через мембраны клетки обеспечивается аквапоринами, которые представляют собой специальные трансмембранные каналы. Аквапорины консервативны во всех клетках — от бактерий до человека, и принадлежат к суперсемейству транспортных белков. У животных аквапорины участвуют во многих физиологических процессах. Так, они обусловливают развитие чувства жажды; участвуют в концентрировании мочи почками, в пищеварении, регуляции температуры тела, секреции и абсорбции спинномозговой жидкости, выделении слез, слюны, пота и желчи, а также в размножении. Например, эпителиальные клетки почек реабсорбируют обратно в кровь 99% воды из первичной мочи, что предотвращает обезвоживание организма. 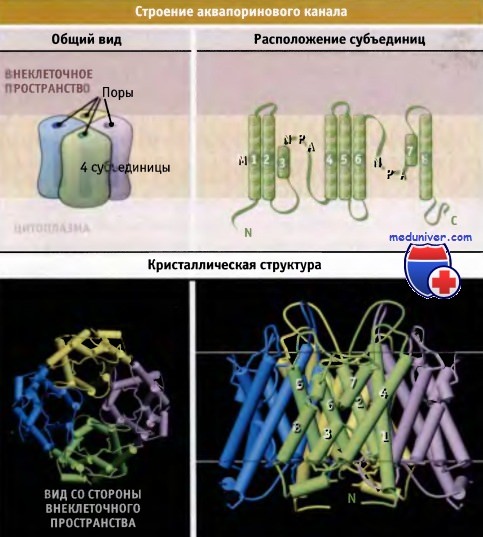 Схематическая структура и кристаллическое строение комплекса аквапоринового канала. Комплекс состоит из четырех идентичных субъединиц, и каждая субъединица образует пору. Кристаллическая структура построена на основании данных Protein Data Bank file 1J4N. Масштаб структуры, видимой со стороны внеклеточного пространства, меньше, чем при виде сбоку. Показано предполагаемое положение мембраны. Если начинается обезвоживание, то клетки нефрона, чувствительные к изменению осмотического давления, обнаруживают повышение осмолярности внеклеточных жидкостей. В ответ стимулируется выброс из гипофиза гормона вазопрессина (антидиуретического гормона). Повышение уровня этого гормона в плазме приводит к уменьшению объема мочи и к ее концентрированию. Связывание вазопрессина с рецепторами клеточной мембраны приводит к быстрой экспрессии аквапорина-2 в апикальной мембране собирательных трубочек почек. Это достигается слиянием внутриклеточных везикул, содержащих аквапорин-2, с апикальной мембраной эпителиальных клеток. Таким образом, в ответ на повышение осмотического градиента, которое наступает при обезвоживании, аквапорины увеличивают реабсорбцию воды из мочи в кровь. Аквапорины являются гомотетрамерными водными каналами, которые регулируют транспорт воды, происходящий за счет осмотического градиента. В отличие от К+-каналов, в которых четыре субъединицы образуют одну пору, в аквапорине каждая субъединица образует независимую пору. Каждая субъединица аквапорина состоит из шести трансмембранных сегментов (M1, М2, М4, М5, М6, и М8), образующих тандемные повторы, каждый из которых, в свою очередь, состоит из трех сегментов. Петли, соединяющие второй и третий трансмембранные сегменты каждого повтора, содержат сигнатурную последовательность аспарагин-пролин-аланин (NPA), одинаковую для всех аквапоринов. Эти две NPA-последовательности в центре водной поры располагаются рядом, образуя часть селективного фильтра. Одна субъединица аквапорина обладает крайне высокой способностью к транспорту воды, которая достигает 3 х 109 молекул в 1 с. В то же время транспорт других растворителей и ионов происходит с существенно меньшей скоростью. Аквапорины содержат трансмембранный водяной канал, через который вода проходит с высокой скоростью и в обоих направлениях. В то же время этот канал не пропускает ионы и даже протоны в форме Н3О+. Такая высокая селективная пропускная способность играет существенную роль в функционировании почек, поскольку одновременная реабсорбция воды и протонов привела бы к несовместимому с жизнью ацидозу. Пора аквапорина подразделяется на три отдела: наружное устье, собственно узкая пора, содержащая селективный фильтр, и внутреннее устье размером 20 А. Эти три отдела формируют транспортный путь, форма которого напоминает часовое стекло. На рисунке ниже выделены остатки аминокислот, участвующие в образовании селективного фильтра. Большая часть стенки канала селективного фильтра построена гидрофобными остатками. Гидрофильные остатки имеют химические группы, необходимые для осуществления селективного транспорта молекул воды. 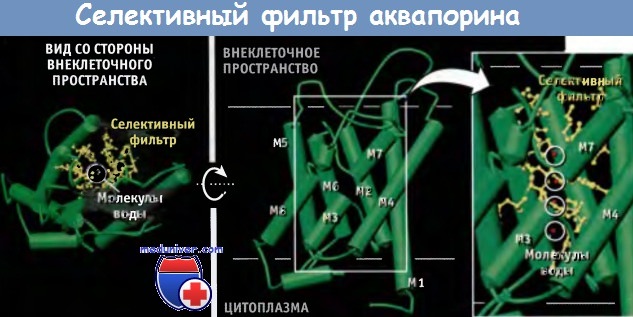 Кристаллическая структура субъединицы аквапоринового канала AQP1. Показано предполагаемое положение мембраны. Остатки, входящие в селективный фильтр, обозначены желтым цветом. Остальная часть белка представлена в виде петель и спиралей (цилиндров). Использованы данные Protein Data Bank file 1J4N. Поры аквапорина характеризуются тремя особенностями, которые обеспечивают им необходимую специфичность по отношению к молекулам воды (рисунок ниже, AQP1 человека): • Ограничения размера. С внешней стороны мембраны в самой узкой точке устье поры достигает 2,8 А в диаметре. Эта область называется область сжатия. Молекулы воды транспортируются через эту область поодиночке, причем гидратированные ионы и протоны в пору не проходят. • Электростатическое отталкивание. Положительный заряд остатков Arg 197, выстилающих пору, способствует электростатическому отталкиванию положительно заряженных молекул и не дает возможность ионам гидрония (Н3О+, образующимся при протонизации молекул воды) проходить через пору. Вместе с тем, в результате внемембранного перекрытия а-спиралей М3 и М7 возникают частичные положительные заряды, которые блокируют протонную проводимость. • Ориентация диполей воды. Селективность увеличивается за счет реориентации диполей воды при образовании водородных связей с частичными положительными зарядами боковых цепей двух аспарагиновых остатков в последовательности NPA (Asn78 и Asn194), находящихся в центре канала. При этом молекулы приобретают определенную ориентацию, и создается второй барьер для входа в пору Н3О+. Способность аквапорина связывать воду приводит к снижению энергетического барьера для процессов транспорта преимущественно по гидрофобному пути. Однако количество актов взаимодействия и величина сродства недостаточны для того, чтобы обеспечить быстрый транспорт воды. Перечисленные особенности селективного фильтра аквапорина обеспечивают быстрый транспорт воды через мембраны, причем протоны, находящиеся в форме Н3О+, и другие ионы в пору не проходят. Различные формы аквапорина играют важную роль в поддержании гомеостаза жидкостей на органном и системном уровнях. В почках экспрессируется несколько изоформ аквапорина, которые участвуют в абсорбции воды из канальцев. Аквапорин I присутствует в эпителиальных клетках проксимальных канальцев и нисходящих тонких участков петли Генле, где обеспечивается постоянная высокая проницаемость для молекул воды. В организме человека аквапорин-1 ежедневно обеспечивает концентрацию 180 л крови до объема 1,5 л, что соответствует объему выводимой мочи. При этом, за счет реабсорбции через апикальную мембрану, около 178,5 л воды из первичного фильтрата возвращается в кровь (реабсорбция воды также происходит внеклеточным путем). У больных с генетическими дефектами аквапорина-1 процесс концентрации мочи нарушен. Экспрессия аквапорина-2 в эпителиальных клетках почек отличается от экспрессии аквапорина-1. Гормон вазопрессин стимулирует экспрессию аквапорина-2 в клетках собирательных трубочек, что приводит к концентрированию мочи. Повышение потребления воды или подавление секреции вазопрессина при потреблении больших количеств алкоголя или кофе приводит к экскреции почками больших количеств разбавленной мочи. У больных несахарным диабетом аквапориновые-2 каналы обладают генетическим дефектом. Такие больные ежедневно выделяют до 20 л мочи. В клетках головного мозга, расположенных рядом с небольшими кровеносными сосудами, экспрессируется аквапорин-4. Он регулирует транспорт воды между паренхимой мозга и сосудами. Аквапорин-4 является потенциальной мишенью для лекарственных средств, используемых при отеке мозга, который часто определяет прогноз при травмах головы и инсульте. Аквапорин-0 экспрессируется только в волокнах хрусталика, и миссенс-мутации приводят к развитию у детей врожденных катаракт. 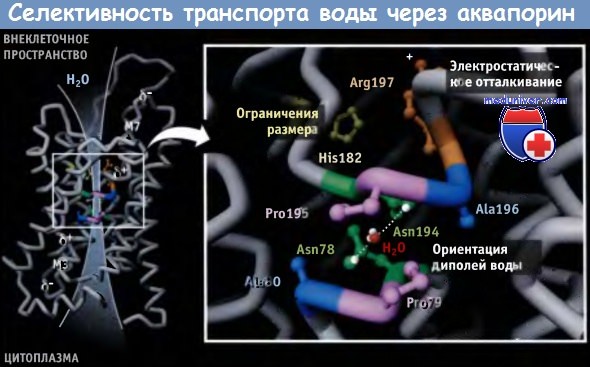 Область поры аквапоринового канала характеризуется тремя особенностями, которые обеспечивают селективный транспорт воды. В рамке показана только одна молекула воды, проникшая в кристаллическую структуру. Атом кислорода этой молекулы образует водородные связи с двумя боковыми цепями Asn. Кристаллическая структура построена на основании данных Protein Data Bank file 1J4N. |