Трудности определения величины риска. Абсолютный и относительный мутационный риск.. Трудности определения величины риска. Трудности определения величины риска. Абсолютный и относительный мутационный риск. Зависимость индукции радиационных мутаций от дозы излучений
 Скачать 153.08 Kb. Скачать 153.08 Kb.
|

|
Трудности определения величины риска. Абсолютный и относительный мутационный риск. Зависимость индукции радиационных мутаций от дозы излучений. Открытие того факта, что между количеством мутаций, возникающих под действием радиации, и дозой радиации имеется прямая зависимость, сыграло очень важную роль в истории радиобиологии и смежных наук, положив начало широкому применению количественных методов исследования. Во многих экспериментах установлена прямая линейная зависимость генетического эффекта излучений от дозы. Известна прямолинейная зависимость количества лизогенных клеток кишечной палочки, в которых профаг переходит в фаг, от дозы рентгеновских лучей. Такого рода связь установлена для многих мутаций у разных организмов. Поэтому принято считать, что частота возникновения генетических эффектов прямо пропорциональна дозе облучения, и дозовая кривая начинается от нулевой точки, т.е. какой бы малой ни была доза радиации, всегда есть риск возникновения мутации. В настоящее время вопрос о действии малых доз радиации в диапазоне от доли рентгена до 10-20 Р тщательно изучается. Что касается разрывов хромосом, то они также появляются в прямолинейной зависимости от дозы. При этом для того, чтобы фрагменты, возникающие при независимых разрывах, могли бы вступить в новые соединения, ведущие к появлению хромосомной перестройки, они должны появиться сравнительно близко к друг другу. Расчёты показывают, что это расстояние не должно превышать 1 мкм. При этом количество хромосомных перестроек, каждая из которых требует для своего возникновения наличия в ядре двух независимых разрывов, в соответствии с теорией вероятности возрастает пропорционально квадрату дозы. Поэтому такая зависимость называется квадратичной, и эффект пропорционален не дозе, как в случае прямолинейной зависимости, а квадрату дозы. Иногда изучаемые эффекты имеют смешанное происхождение – например, рецессивные мутации могут быть и точковыми мутациями, и микроаберрациями. В этом случае используют линейно- квадратичную модель. Линейная беспороговая модель очень простая и обычно используется для вычисления радиационной опасности. Математические она выражается следующим образом: Риск = α× Доза, где α – константа. Квадратичная модель описывается формулой: Риск =α× Доза× Доза, где α – константа. Линейно- квадратичная модель описывается уравнением следующего типа: Число трансформированных клеток = α× Доза + β × Доза× Доза, где α и β – константы. 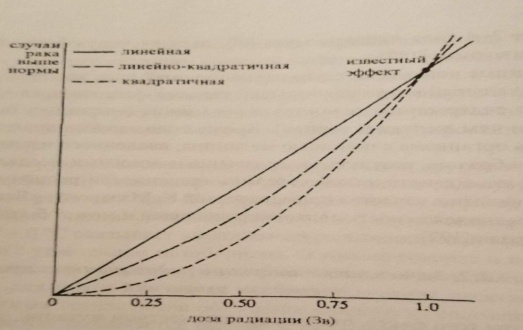 Конечно, основываясь на результатах изучения клеток, очень трудно делать выводы о том, какой формы должна быть дозовая кривая при облучении в малых дозах. Если мы используем линейную модель в том случае, когда правильно является линейно-квадратичная, мы переоценим риск. И наоборот, если используем линейно-квадратичную модель, когда нам следовало бы использовать линейную, мы недооцениваем риск. Очень важно понимать, что мы не можем в настоящее время доказать, какая модель правильная. Существует ещё один усложняющий фактор. При высоких дозах радиации возрастает гибель клеток, при этом происходит элиминация клеток с мутациями. Таким образом, кривая доза-эффект при высоких дозах может «загибаться вниз». Эта кривая, характерная для радиационно-генетических эффектов, приводит к парадоксальным выводам - с увеличением дозы облучения (при дозах, приближающихся к летальным) частота мутаций уменьшается. Это явление получило название «аномальной зависимости в области высоких доз». И в этом заключается основное отличие радиационно-генетических эффектов от общебиологических, для которых характерно с увеличением дозы только возрастание эффекта. 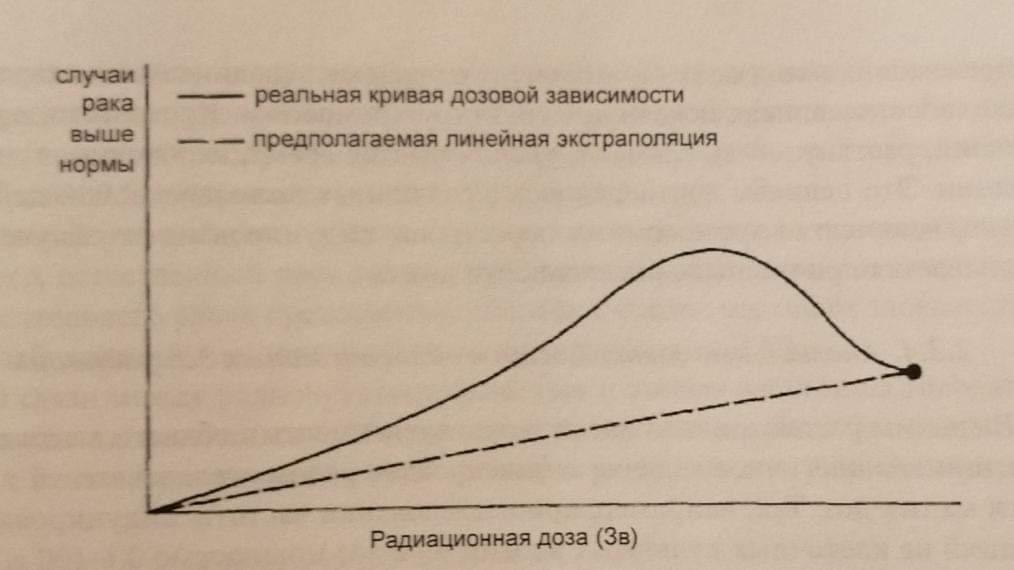 Эффект мощности дозы. Помимо проблемы экстраполяции с высоких доз на низкие, существуют еще факторы, усложняющие вычисление генетического риска, обусловленного облучением в малых дозах. При воздействии радиации та же доза, растянутая во времени (при низкой мощности дозы), может быть менее опасной, чем при остром облучении, вследствие способности клеток препарировать повреждения ДНК. Это можно сопоставить с известными фактами, что, например, приём 100 мг стрихнина или мышьяка, или потери 10 л крови гораздо более опасны, когда всё это происходит в несколько секунд, чем когда это растянуто на месяцы. Этот аргумент применим и к воздействию радиации. Поскольку большинство хромосомных перестроек зависят в своём появлении от двух независимых разрывов, на их образование влияет мощность дозы радиации. В случае, если данная доза дается сразу за короткое время, мы можем ожидать максимальный эффект для всех возможных перекомбинаций фрагментов, возникающих одновременно. Если же давать ту же дозу за длительный период времени или по частям, то многие разрывы, возникая в разное время, не смогут вступить в перекомбинацию и поэтому фрагмент от разрыва соединяется в старом порядке, восстанавливая исходную структуру хромосомы. Кроме того, при облучении, растянутым во времени, увеличивается время, необходимое для репарации. Вывод: количество хромосомных перестроек, индуцированных облучением, уменьшается при уменьшении мощности дозы. Аномальные зависимости в области малых доз радиации. Выше мы рассматривали аномальную зависимость в области высоких доз радиации. Аналогичное явление зафиксировано рядом исследователей и в области малых доз. Так, например, при исследовании частоты индуцированных мутаций на клеточных культурах в диапазоне доз мощностью 0,1-100 сГр обнаружена нелинейная зависимость эффекта от дозы. Так, при мощности экспозиционной дозы от 0,1 до 1 сГр наблюдали эффект обратной зависимости от дозы, а при мощности 1-100 сГр обнаружили прямую зависимость. Считают, что появление на кривой зависимости доза-эффект участка с минимальным уровнем мутаций отражает процесс активации внутриклеточных механизмов, защищающих клетку от неблагоприятного действия мутагенных факторов. Данные механизмы обеспечивают контроль клеточного цикла, детоксикацию активных форм кислорода и репарацию генетических повреждений. По-видимому, именно реализация того или иного клеточного механизма отражается на характере зависимости эффекта от дозы и показывает специфику эффектов в диапазоне малых доз. Возможно, что одной из причин «обратной зависимости» в области малых доз является «байстэндер» эффект (заключается в передаче радиационных сигналов от облученных клеток необлученным). Модели абсолютного и относительного риска. Второй усложняющий фактор связан с тем, что генетические эффекты зависят не только от величины и мощности дозы радиации, но и от уровня спонтанных или естественных мутаций. Необходимо различать модель абсолютного риска, в которой принимается, что риск, вызванный обучением, увеличивается на определенную величину, и модель относительного риска, в которой принимается, что риск возрастает на величину, пропорциональную естественному риску. Частоту индуцированных мутаций можно выразить в абсолютных терминах (абсолютный мутационный риск - это вероятность мутирования данного гена при облучении в 1 Зв) или в относительных единицах - отношение такое частоты мутаций на зиверт к частоте спонтанных мутаций. Это отношение называется относительным мутационным риском и выражается в Зв-1. Часто используют показатель удваивающей дозы, т.е. дозы, которая вызовет 100 %-ное увеличение частоты спонтанных мутаций. Удваивающая доза - это величина, обратная относительно мутационную риску. Предположим, например, что 1 Зв увеличивает уровень мутаций на 50% сверх спонтанного уровня. Если 50% выразить в долях, получим 0,5. Таким образом, относительный мутационный риск равен 0,5, а удваивающая доза составит 1: 0,5 или 2 Зв. Относительный риск в ряде случаев является важным показателем. Например, естественный риск рака увеличивается с возрастом, поэтому модель относительного риска предсказывает большее число смертей от злокачественных болезней, чем модель абсолютного риска.  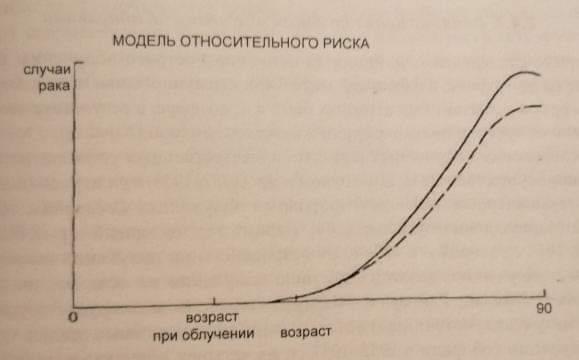 Что усложняет определение величины генетического риска ? Генетические эффекты зависят от величины (влияние больших и малых доз) и мощности дозы радиации и от уровня спонтанных / естественных мутаций. Какой формы должна быть дозовая кривая при облучении в малых дозах? Если мы используем линейную модель в том случае, когда правильно является линейно-квадратичная, мы переоценим риск. И наоборот, если используем линейно-квадратичную модель, когда нам следовало бы использовать линейную, мы недооцениваем риск. Существует ли зависимость числа мутаций в хромосомах от мощности дозы? Количество хромосомных перестроек, индуцированных облучением, уменьшается при уменьшении мощности дозы. Какое основное отличие радиационно-генетических эффектов от общебиологических? С увеличением дозы облучения частота мутаций уменьшается. В каких случаях используют линейно-квадратичную модель? Когда изучаемые эффекты имеют смешанное происхождение – например, рецессивные мутации могут быть точковыми мутациями. Есть ли причина «обратной зависимости» в области малых доз? Одной из причин является «байстэндер» эффект (заключается в передаче радиационных сигналов от облученных клеток необлученным). |