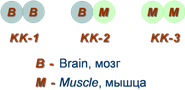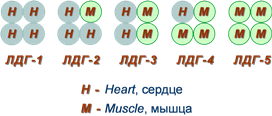Занятие 4. Ферменты. Биологическая роль. Механизм и особенности ферментативного катализа. Кофакторы и коферменты
 Скачать 345.26 Kb. Скачать 345.26 Kb.
|

|
Занятие №4 ТЕМА: ФЕРМЕНТЫ. БИОЛОГИЧЕСКАЯ РОЛЬ. МЕХАНИЗМ И ОСОБЕННОСТИ ФЕРМЕНТАТИВНОГО КАТАЛИЗА. КОФАКТОРЫ И КОФЕРМЕНТЫ. Цель: Рассмотреть современные представления о строении ферментов, их биологической роли и особенностях ферментативного катализа. Составить представление о механизме действия ферментов, изучить строение и участие в метаболизме кофакторов и коферментов. 1. Ферменты, определение. Биологическая роль ферментов. Понятие апофермент, кофермент, субстрат, продукт реакции, активаторы и ингибиторы ферментов. Ферме́нты или энзи́мы — обычно достаточно сложные молекулы белка, рибозимы или их комплексы, ускоряющие химические реакции в живых системах. Каждый фермент, свернутый в определённую структуру, ускоряет соответствующую химическую реакцию: реагенты в такой реакции называются субстратами, а получающиеся вещества — продуктами. Ферменты специфичны к субстратам: АТФ-аза катализирует расщепление только АТФ, а киназа фосфорилазы фосфорилирует только фосфорилазу. Ферментативная активность может регулироваться активаторами (повышаться) и ингибиторами (понижаться). Белковые ферменты синтезируются на рибосомах, а РНК — в ядре. Наука о ферментах называется энзимологией, а не ферментологией (чтобы не смешивать корни слов латинского и греческого языков). Биологическая роль ферментов. Ускоряет реакции в клетках, ферменты катализируют 2 тыс-3 тыс реакций обмена, есть так же вовлеченные в передачу сигнала, процессе дыхания, мышечного сокращения, свертываемость крови, транспорт веществ, обезвреживание токсичных и чужеродных соединений, нейротрансмиссия. 1.Простетические группы прочно связаны с белками и даже могут быть присоединены ковалентными связями. Часто играют важную роль в функционировании ферментов. Белок без простетической группы называется «апобелок», а белок с присоединенной группой — «холобелок» (или, соответственно, в случае ферментов — апофермент и холофермент). Примером может являться гем, который является простетической группой в молекуле гемоглобина. 2.Кофактор -небелковое вещество, которое обязательно должно присутствовать в организме в небольших количествах, чтобы соответствующие ферменты смогли выполнить свои функции. В состав кофактора входят коферменты и ионы металлов (например, ионы натрия и калия). 3.Субстрат -вещество, подвергающееся превращению под действием фермента; 4. Активаторы – вещества, которые повышают скорость ферментативных реакций, увеличивают активность ферментов. Они бывают органической и неорганической природы. Активаторы органической природы: желчные кислоты (активируют поджелудочную ли пазу), энтерокиназа (активирует трипсиноген), глутатион, цистеин, витамин С (повышают активность оскидоредуктаз). Активаторы неорганической природы: например, HCl активирует пепсиноген, ионы ме таллов (Na, Cl, K, Mg, Mn, Zn) активируют очень многие ферменты. Ионы металлов: а) спо собствуют образованию ферментсубстратного комплекса; б) служат донорами и акцептора ми электронов; в) принимают участие в образовании активного центра ферментов (Zn в со ставе карбангидразы, Fe – в составе цитохромов, каталазы, пероксидазы); г) выступают в ро ли аллостерических регуляторов. 5. Ингибиторы – вещества, которые уменьшают активность ферментов и замедляют химические реакции. Различают обратимое и необратимое ингибирование: Если ингибитор связывается с молекулой фермента слабыми связями (Е+И ↔ ЕИ) то такой ингибитор легко удаляется и активность фермента восстанавливается; Если ингибитор связывается с молекулой фермента прочными ковалентными связями (Е+И→ ЕИ), то наступает необратимое подавление активности фермента 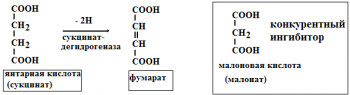 Необратимое ингибирование происходит при денатурация ферментовбелков под действием концентрированных кислот и щелочей, солей тяжелых металлов, ультрафиолетовом облучении. Некоторые ингибиторы образуют прочные недиссоциируемые связи с функцио нальными группами в активных центрах ферментов. Например, цианиды связываются с же лезом в ферментахгемопротеинах. Фосфорорганические яды (табун, зарин, Vгазы) образу ют прочные связи с остатками серина и треонина входящими в состав многих ферментов. Обратимое ингибирование делится на конкурентное и неконкурентное. Конкурентное ин гибирование вызывается веществами, структурно сходными с субстратом и взаимодейст вующими с активным центром фермента. Например, малоновая кислота, является конку рентным ингибитором сукцинатдегидрогеназы, посколььку похожа на янтарную кислоту (также имеет 2 карбоксильных группы). Поэтому, малоновая кислота легко связывается с ак тивным центром сукцинатдегидрогеназы, вытесняя оттуда субстрат – янтарную кислоту. Од нако, фермент неспособен это сделать с малоновой кислотой, которая короче на 1 атом углерода. Поэто му если прибавить ма лоновую кислоту в концентрации, превышающей концентрацию янтарной кислоты, то реакция прекратится, поскольку малонат за блокирует активный центр сукцинатдегидрогеназы Конкурентные ингибиторы нередко используются в качестве лекарственных средств. Например, антимикробные препараты сульфаниламиды являются структурными аналогами парааминобензойной кислоты из которой микроорганизмы синтезируют необходимый им для размножение витамин В9 (фолиевую кислоту). Многие антибиотики конкурентно тормо зят синтез белка микроорганизмами или репликацию ДНК. Потивоопухолевые препараты (метотрексат, антагонист витамина В9) блокирует репликацию ДНК в опухолевых клетках. Неконкурентные ингибиторы не имеют структурного сходства к субстрату и при соединяются не к активному центру, а к другим участкам, в том числе и к аллостерическому центру. Ингибирование происходит вследствие разрушения или необратимой химической модификации функциональных групп ферментов. Примеры: а) алкилирующие агенты (йодацетамид) необратимо реагируют с SН–группами ферментов Е–SH + ICH2CОNH2 → E–SCH2 –CОNH2 + HI (фермент) (йодацетамид) комплекс ферментингибитор б) препараты ФОС (фосфорорганических соединений) это высокотоксичные яды для насеко мых и теплокровных животных. Они взаимодействуют с гидроксигруппой серина в активном центре фермента ацетилхолинэстеразы: 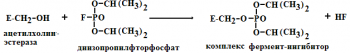 в) тетурам – ингибитор ацетальдегиддегидрогеназы (используют при лечении алкоголизма). Согласно общепринятым представлениям о механизме простейших ферментативных реакций, которые были предложены Михаэлисом и Меитеи, субстрат S и фермент Е находятся в равновесии с очень быстро образующимся фермент-субстратным комплексом ES. Этот комплекс сравнительно медленно распадается на продукт реакции Р и свободный фермент Е. Таким образом, стадия распада фермент-субстратного комплекса на продукты реакции является лимитирующей. где кj константа скорости образования фермент-субстратного комплекса; к— константа скорости обратной реакции, распада фермент-субстратного комплекса; к2 — константа скорости образования продукта реакции. 2. Классификация и номенклатура ферментов. Современные классификация и номенклатура ферментов были разработаны Комиссией по ферментам Международного биохимического союза и утверждены на V Международном биохимическом конгрессе в 1961 г. в Москве. Необходимость систематики номенклатуры диктовалась прежде всего стремительным ростом числа вновь открываемых ферментов, которым разные исследователи присваивали названия по своему усмотрению. Более того, одному и тому же ферменту часто давали два или несколько названий, что вносило путаницу в номенклатуру. Некоторые названия ферментов вообще не отражали тип катализируемой реакции, а при наименовании фермента исходили из названия субстрата, на который действует фермент, с добавлением окончания -аза: в частности, амилазы(ферменты, гидро-лизирующие углеводы), липазы (действующие на липиды), протеиназы (гидролизирующие белки) и т.д. До 1961 г. не было и единой классификации ферментов. Трудности заключались в том, что разные исследователи за основу классификации ферментов брали различные принципы. Комиссией были рассмотрены 3 принципа, которые могли служить основой для классификацииферментов и их обозначения. Первый принцип – химическая природа фермента, т.е. принадлежность к флавопротеинам, пиридоксальфосфатпротеинам, гемо-протеинам, металлопротеинам и т. д. Однако этот принцип не мог служить общей основой для классификации, так как только для небольшого числа ферментов известны простетические группы, доступные идентификации и прямому определению. Второй принцип – химическая природа субстрата, на который действует фермент. По этому принципу трудно классифицировать фермент, так как в качестве субстрата могут служить разнообразные соединения внутри определенного класса веществ (белки, углеводы, липиды, нуклеиновые кислоты) и бесчисленное множество промежуточных продуктов обмена. В основу принятой классификации положен третий принцип – тип катализируемой реакции , который является специфичным для действия любого фермента. Этот принцип логично использовать в качестве основы для классификации и номенклатуры ферментов. Таким образом, тип катализируемой химической реакции в сочетании с названием субстрата(субстратов) служит основой для систематического наименования ферментов. Согласно Международной классификации, ферменты делят на шесть главных классов, в каждом из которых несколько подклассов: 1) оксидоредуктазы; 2) трансферазы; 3) гидролазы; 4) лиазы; 5) изомеразы; 6) лигазы (синтетазы) (табл. 4.5). Оксидоредуктазы. К классу оксидоредуктаз относят ферменты, катализирующие с участием двух субстратов окислительно-восстановительные реакции, лежащие в основе биологического окисления. Систематические названия их составляют по форме «донор: акцептор оксидоредуктаза». Например, лактат: НАД+ оксидоредуктаза для лактатдегидрогеназы (ЛДГ). Различают следующие основные оксидоредуктазы: аэробные дегидро-геназы или оксидазы, катализирующие перенос протонов (электронов) непосредственно на кислород; анаэробные дегидрогеназы, ускоряющие перенос протонов (электронов) на промежуточный субстрат, но не на кислород; цитохромы, катализирующие перенос только электронов. К этому классу относят также гемсодержащие ферменты каталазу и пероксидазу, катализирующие реакции с участием перекиси водорода. Трансферазы. К классу трансфераз относят ферменты, катализирующие реакциимежмолекулярного переноса различных атомов, групп атомов и радикалов. Наименование их составляется по форме «донор: транспортируемая группа – трансфераза». Различают трансферазы, катализирующие перенос одноуглеродных остатков, ацильных, гликозильных, альдегидных или кетонных, нуклеотидных  остатков, азотистых групп, остатков фосфорной и серной кислот и др. Например: метил- и формилтрансферазы, ацетилтрансферазы, амино-трансферазы, фосфотрансферазы и др. Гидролазы. В класс гидролаз входит большая группа ферментов, катализирующих расщепление внутримолекулярных связей органических веществ при участии молекулы воды. Наименование их составляют по форме «субстрат-гидролаза». К ним относятся: зстеразы – ферменты, катализирующие реакции гидролиза и синтеза сложных эфиров; гликозидазы, ускоряющие разрыв гликозидных связей; фосфатазы и пептидгидролазы, катализирующие гидролиз фосфоангидридных и пептидных связей; ами-дазы, ускоряющие разрыв амидных связей, отличных от пептидных, и др. Лиазы. К классу лиаз относят ферменты, катализирующие разрыв связей С—О, С—С, С—N и других, а также обратимые реакции отщепления различных групп от субстратов не гидролитическим путем. Эти реакции сопровождаются образованием двойной связи или присоединением групп к месту разрыва двойной связи. Ферменты обозначают термином «субстрат-лиазы». Например, фумарат-гидратаза (систематическое название «L-малат-гидролаза») катализирует обратимое отщепление молекулы воды от яблочной кислоты с образованием фумаровой кислоты. В эту же группу входят декарбоксилазы (карбокси-лиазы), амидин-лиазы и др. Изомеразы. К классу изомераз относят ферменты, катализирующие взаимопревращения оптических и геометрических изомеров. Систематическое название их составляют с учетом типареакции: «субстрат – цис-транс-изомераза». Если изомеризация включает внутримолекулярный перенос группы, фермент получает название «мутаза». К этому же классу относят рацемазы и эпимеразы, действующие на амино- и оксикислоты,углеводы и их производные; внутримолекулярные оксидоредуктазы, катализирующие взаимопревращения альдоз и кетоз; внутримолекулярные трансферазы, переносящие ацильные, фосфорильные и другие группы, и т.д. Лигазы (синтетазы). К классу лигаз относят ферменты, катализирующие синтез органическихвеществ из двух исходных молекул с использованием энергии распада АТФ (или другого нуклеозидтрифосфата). Систематическое название их составляют по форме «X : Y лигаза», где X и Y обозначают исходные вещества. В качестве примера можно назвать L-глутамат: аммиаклигазу (рекомендуемое сокращенное название «глутаминсинтета-за»), при участии которой из глутаминовой кислоты и аммиака в присутствии АТФ синтезируется глутамин. 3. Строение ферментов. Активный центр ферментов, состав, формирование, роль. Функциональные группы аминокислот, входящих в его состав. Давно выяснено, что все ферменты являются белками и обладают всеми свойствами белков. Поэтому подобно белкам ферменты делятся на простые и сложные. Простые ферменты состоят только из аминокислот – например, пепсин , трипсин, лизоцим. Сложные ферменты (холоферменты) имеют в своем составе белковую часть, состоящую из аминокислот – апофермент, и небелковую часть – кофактор. Примером сложных ферментовявляются сукцинатдегидрогеназа (содержит ФАД), аминотрансферазы (содержат пиридоксальфосфат), пероксидаза (содержит гем), лактатдегидрогеназа (содержит Zn2+), амилаза (содержит Ca2+). Кофактор, в свою очередь, может называться коферментом (НАД+, НАДФ+, ФМН, ФАД, биотин) или простетической группой (гем, олигосахариды, ионы металлов Fe2+, Mg2+, Ca2+, Zn2+). Деление на коферменты и простетические группы не всегда однозначно: • если связь кофактора с белком прочная, то в этом случае говорят о наличии простетической группы, • но если в качестве кофактора выступает производное витамина - то его называют коферментом, независимо от прочности связи. Для осуществления катализа необходим полноценный комплекс апобелка и кофактора, по отдельности катализ они осуществить не могут. Кофактор входит в состав активного центра, участвует в связывании субстрата или в его превращении. Как многие белки, ферменты могут быть мономерами, т.е. состоять из одной субъединицы, и полимерами, состоящими из нескольких субъединиц. Структурно-функциональная организация ферментов В составе фермента выделяют области, выполняющие различную функцию: 1. Активный центр – комбинация аминокислотных остатков (обычно 12-16), обеспечивающая непосредственное связывание с молекулой субстрата и осуществляющая катализ. Аминокислотные радикалы в активном центре могут находиться в любом сочетании, при этом рядом располагаются аминокислоты, значительно удаленные друг от друга в линейной цепи. В активном центре выделяют два участка: якорный (контактный, связывающий) – отвечает за связывание и ориентацию субстрата в активном центре, каталитический – непосредственно отвечает за осуществление реакции. 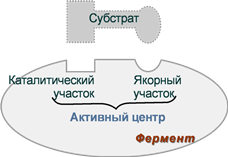 Схема строения ферментов У ферментов, имеющих в своем составе несколько мономеров, может быть несколько активных центров по числу субъединиц. Также две и более субъединицы могут формировать один активный центр. У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора.  Схема формирования сложного фермента 2. Аллостерический центр (allos – чужой) – центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Связывание с аллостерическим центром какой-либо молекулы (называемой активатором или ингибитором, а также эффектором, модулятором, регулятором) вызывает изменение конфигурации белка-фермента и, как следствие, скорости ферментативной реакции. Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся в разных субъединицах. 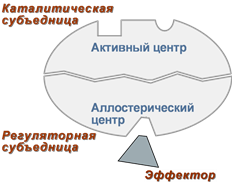 Схема строения аллостерического фермента В качестве такого регулятора может выступать продукт данной или одной из последующих реакций, субстрат реакции или иное вещество (см "Регуляция активности ферментов"). Изоферменты Изоферменты – это молекулярные формы одного и того же фермента, возникшие в результате небольших генетических различий в первичной структуре фермента, но катализирующие одну и ту же реакцию. Изоферменты отличаются сродством к субстрату, максимальной скоростью катализируемой реакции, чувствительностью к ингибиторам и активаторам, условиями работы (оптимум pH и температуры). Как правило, изоферменты имеют четвертичную структуру, т.е. состоят из двух или более субъединиц. Например, димерный фермент креатинкиназа (КК) представлен тремя изоферментными формами, составленными из двух типов субъединиц: M (англ. muscle – мышца) и B (англ. brain – мозг). Креатинкиназа-1 (КК-1) состоит из субъединиц типа B и локализуется в головном мозге, креатинкиназа-2 (КК-2) – по одной М- и В-субъединице, активна в миокарде, креатинкиназа-3 (КК-3) содержит две М-субъединицы, специфична для скелетной мышцы.
Также существует пять изоферментов лактатдегидрогеназы (роль ЛДГ) – фермента, участвующего в обмене глюкозы. Отличия между ними заключаются в разном соотношении субъединиц Н (англ. heart – сердце) и М (англ. muscle – мышца). Лактатдегидрогеназы типов 1 (Н4) и 2 (H3M1) присутствуют в тканях с аэробным обменом (миокард, мозг, корковый слой почек), обладают высоким сродством к молочной кислоте (лактату) и превращают его в пируват. ЛДГ-4 (H1M3) и ЛДГ-5 (М4) находятся в тканях, склонных к анаэробному обмену (печень, скелетные мышцы, кожа, мозговой слой почек), обладают низким сродством к лактату и катализируют превращение пирувата в лактат. В тканях с промежуточным типом обмена (селезенка, поджелудочная железа, надпочечники, лимфатические узлы) преобладает ЛДГ-3 (H2M2). Еще одним примером изоферментов является группа гексокиназ, которые присоединяют фосфатную группу к моносахаридам гексозам и вовлекают их в реакции клеточного метаболизма. Из четырех изоферментов выделяется гексокиназа IV (глюкокиназа), которая отличается от остальных изоферментов высокой специфичностью к глюкозе, низким сродством к ней и нечувствительностью к ингибированию продуктом реакции. Мультиферментные комплексы В мультиферментном комплексе несколько ферментов прочно связаны между собой в единый комплекс и осуществляют ряд последовательных реакций, в которых продукт реакции непосредственно передается на следующий фермент и является только его субстратом. Возникает туннельный эффект, т.е. субстрат попадает в созданный ферментами "туннель". В результате промежуточные метаболиты избегают контакта с окружающей средой, снижается время их перехода к следующему активному центру и значительно ускоряется скорость реакции. 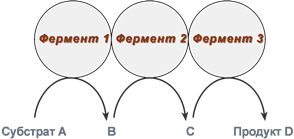 Строение мульферментного комплекса Например, пируватдегидрогеназный комплекс (пируватдегидрогеназа), превращающий пируват в ацетил-SКоА, α-кетоглутаратдегидрогеназный комплекс (в цикле трикарбоновых кислот) превращающий α-кетоглутарат в сукцинил-SКоА, комплекс под названием "синтаза жирных кислот" (или пальмитатсинтаза), синтезирующий пальмитиновую кислоту карбамоилфосфатсинтетаза, первый фермент синтеза мочевины в печени и синтеза пиримидиновых оснований Абзимы Абзимами называются антитела, имеющие каталитическую функцию (англ. abzymes, antibodiesas enzymes) и катализирующие конкретные реакции. Такая способность возникает в результате формирования промежуточного продукта при связывании антитела с антигеном (имитация переходного комплекса E-X ферментативной реакции). 4. Особенности ферментативного катализа. Виды специфичности. Ферментативный катализ (биокатализ), ускорение биохимических реакций при участии белковых макромолекул. называемых ферментами (энзимами). Ферментативный катализ - разновидность катализа, хотя термин "ферментация" (брожение)известен с давних времен, когда еще не было понятия химического катализа. Важнейшие особенности ферментативного катализа - эффективность, специфичность и чувствительность к регуляторным воздействиям. Ферменты увеличивают скорость химического превращения субстрата по сравнению с неферментативной реакцией в 109-1012 раз. Столь высокая эффективность обусловлена особенностями строения активного центра. Принято считать, что активный центр комплементарен переходному состоянию субстрата при превращении его в продукт. Благодаря этому стабилизируется переходное состояние и понижается активационный барьер реакции. В ферментативной реакции можно выделить следующие этапы: 1. Присоединение субстрата (S) к ферменту (E) с образованием фермент-субстратного комплекса (E-S). 2. Преобразование фермент-субстратного комплекса в один или несколько переходных комплексов (E-X) за одну или несколько стадий. 3. Превращение переходного комплекса в комплекс фермент-продукт (E-P). 4. Отделение конечных продуктов от фермента. Ферменты обладают более высокой специфичностью действия по сравнению с неорганическими катализаторами. Различают специфичность по отношению к типу химической реакции, катализируемой ферментом, и специфичность по отношению к субстрату. Эти два вида специфичности характерны для каждого фермента. Специфичность по отношению к субстрату – это предпочтительность фермента к субстрату определенной структуры в сравнении с другими субстратами. Различают 4 вида субстратной специфичности ферментов: 1. Абсолютная специфичность – способность фермента катализировать превращение только одного субстрата. Например – глюкокиназа фосфорилирует только глюкозу, аргиназа расщепляет только аргинин, уреаза – мочевину. 2. Относительная специфичность – фермент катализирует превращение нескольких субстратов, имеющих один тип связи. Например – липаза расщепляет сложноэфирную связь в триацилглицеролах. 3. Относительная групповая специфичность – фермент катализирует превращение нескольких субстратов, имеющих один тип связи, но требуется наличие определенных функциональных групп, входящих в состав субстратов. Например, все протеолитические ферменты расщепляют пептидную связь, но пепсин – образованную аминогруппами ароматических аминокислот, химотрипсин – образованную карбоксильными группами этих же аминокислот, трипсин – пептидную связь, образованную карбоксильной группой лизина, аргинина. 4. Стереохимическая специфичность – фермент катализирует превращение только одного стереоизомера. Например, бактериальная аспартатдекарбоксилаза катализирует декарбоксилирование только L-аспартата и не действует на D-аспарагиновую кислоту. Каталитическая специфичность Фермент катализирует превращение присоединённого субстрата по одному из возможных путей его превращения. Это свойство обеспе-чивается строением каталитического участка активного центра фермента и называется ката-литической специфичностью, или специфич-ностью пути превращения субстрата. Так, молекула глюкозо-6-фосфата в клетках печени человека — субстрат 4 различных ферментов: фосфоглюкомугазы, глюкозо-6-фосфатфосфатазы, фосфоглюкоизомеразы и глюкозо-6-фосфатдегидрогеназы. Однако из-за особенностей строения каталитических участков этих ферментов происходит рахчичное превращение этого соединения с образованием 4 различных продуктов (см. схему ниже). 5. Механизм действия ферментов. Этапы катализаВ ферментативной реакции можно выделить следующие этапы: 1. Присоединение субстрата (S) к ферменту (E) с образованием фермент-субстратного комплекса (E-S). 2. Преобразование фермент-субстратного комплекса в один или несколько переходных комплексов (E-X) за одну или несколько стадий. 3. Превращение переходного комплекса в комплекс фермент-продукт (E-P). 4. Отделение конечных продуктов от фермента. Механизмы катализа
1. Кислотно-основной катализ – в активном центре фермента находятся группы специфичных аминокислотных остатков, которые являются хорошими донорами или акцепторами протонов. Такие группы представляют собой мощные катализаторы многих органических реакций. 2. Ковалентный катализ – ферменты реагируют со своими субстратами, образуя при помощи ковалентных связей очень нестабильные фермент-субстратные комплексы, из которых в ходе внутримолекулярных перестроек образуются продукты реакции. Типы ферментативных реакций1. Тип "пинг-понг" – фермент сначала взаимодействует с субстратом А, отбирая у него какие либо химические группы и превращая в соответствующий продукт. Затем к ферменту присоединяется субстрат В, получающий эти химические группы. Примером являются реакции переноса аминогрупп от аминокислот на кетокислоты - трансаминирование. 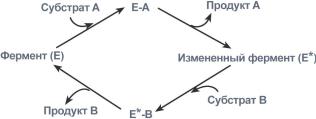 Ферментативная реакция по типу "пинг-понг"2. Тип последовательных реакций – к ферменту последовательно присоединяются субстраты А и В, образуя "тройной комплекс", после чего осуществляется катализ. Продукты реакции также последовательно отщепляются от фермента. 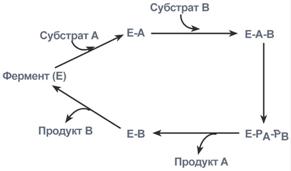 Ферментативная реакция по типу "последовательных реакций"3. Тип случайных взаимодействий – субстраты А и В присоединяются к ферменту в любом порядке, неупорядоченно, и после катализа так же отщепляются. 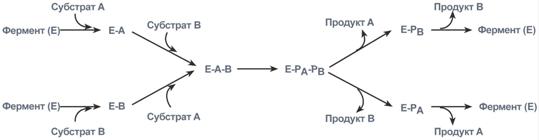 6. Кофакторы ферментов: ионы металлов (на примере карбоксипептидазы А, амилазы) и нуклеотидные кофакторы: УТФ, ЦТФ, ГТФ, АТФ. Кофактор -небелковое вещество, которое обязательно должно присутствовать в организме в небольших количествах, чтобы соответствующие ферменты смогли выполнить свои функции. В состав кофактора входят коферменты и ионы металлов (например, ионы натрия и калия). Гексокиназа катализирует перенос концевого, у-фосфатного остатка молекулы АТФ на глюкозу с образованием глюкозо-6-фосфата: 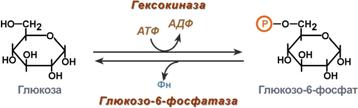 Участие ионов магния в присоединении субстрата в активном центре гексокиназы. В активном центре гексокиназы есть участки связывания для молекулы глюкозы и комплекса Мg2 +-АТФ. В результате ферментативной реакции происходит перенос концевого, gamma-фосфатного остатка молекулы АТФ на глюкозу с образованием глюкозо-6-фосфата.  Гексокиназа - первый фермент метаболизма глюкозы Наличие глюкозы в клетке обеспечивается, в первую очередь, облегченной диффузией ее из крови в цитозоль при участии специальных транспортных белков - глюкозных транспортеров(ГлюТ). Активация глюкозы После перемещения через мембраны глюкоза в цитозоле немедленно фосфорилируется ферментом гексокиназой, в связи с чем фермент образно называют "ловушка глюкозы". Фосфорилирование глюкозы решает несколько задач: фосфатный эфир глюкозы не в состоянии выйти из клетки, так как молекула отрицательно заряжена и отталкивается от фосфолипидной поверхности мембраны, наличие заряженной группы обеспечивает правильную ориентацию молекулы в активном центре фермента уменьшается концентрация свободной (нефосфорилированной) глюкозы, что способствует диффузии новых молекул из крови. Дефосфорилирование глюкозы осуществляется глюкозо-6-фосфатазой. Этот фермент есть только в печени и почках. В эпителии канальцев почек работа фермента связана с реабсорбцией глюкозы. В гепатоцитах фермент необходим, когда печень поддерживает гомеостаз глюкозы в крови. Реакции фосфорилирования и дефосфорилирования глюкозы Кроме внешней среды (крови) источником глюкозы для клеток являются запасы гликогена, который используется как внутриклеточный резерв. Вместе с этим, гепатоциты и канальцевый эпителий почек обладают способностью синтезировать глюкозу из неуглеводных компонентов (глюконеогенез). Но в данном случае глюкоза уходит из этих клеток в кровь и используется другими клетками. Особенности глюкокиназы Существуют принципиальные отличия метаболизма глюкозы в печени от других тканей. Это объясняется рядом причин и, в частности, наличием в тканях различных изоферментов гексокиназы. Для печени характерен особый изофермент гексокиназа IV, получивший собственное название – глюкокиназа. Отличиями этого фермента от гексокиназ других тканей являются: низкое сродство к глюкозе (в 1000 раз меньше), что ведет к захвату глюкозы печенью только при ее высокой концентрации в крови (после еды), продукт реакции глюкозо-6-фосфат не ингибирует фермент, в то время как в других тканях гексокиназа чувствительна к такому влиянию. Это позволяет гепатоциту в единицу времени захватывать глюкозы больше, чем он может сразу же утилизовать, чувствительность к действию инсулина – фермент активируется этим гормоном. Благодаря таким отличиям гепатоцит может эффективно захватывать глюкозу после еды, накапливать и впоследствии метаболизировать ее в любом направлении – синтез гликогена, пентозофосфатный путь, окисление до ацетил-SКоА и синтез липидов. 7. Коферментные функции витаминов (на примере трансаминаз и дегидрогеназ, витаминов В6; РР; В2). Витамины играют важную роль в обмене веществ. В настоящее время известны не только те реакции, для нормального течения которых необходим тот или иной витамин, но и ферменты, в со став коферментов которых входят витамины (табл. 14). Описано более 100 таких ферментов. Недостаточное поступление витаминов с пищей, нарушение их всасывания и усвоения, повышенная потребность организма в них могут приводить к специфическим для каждого витамина наруше ниям обмена веществ и физиологических функций, снижению ра ботоспособности. Длительный дефицит поступления витаминов вызывает специфические заболевания (гиповитаминозы и авитаминозы).
8. Структура и биологическая роль коферментов: ТПФ, НАД и НАДФ, ФАД и ФМН, ПФ, биотина, ТГФК, КоА, (аскорбиновая кислота для стомат факультета) Участие коферментов в метаболизме. Коферменты в каталитических реакциях осуществляют транспорт различных групп атомов, электронов или протонов. Коферменты связываются с ферментами: - ковалентными связями; - ионными связями; - гидрофобными взаимодействиями и т.д. Один кофермент может быть коферментом для нескольких ферментов. Многие коферменты являются полифункциональными (например, НАД, ПФ). В зависимости от апофермента зависит специфичность холофермента. Все коферменты делят на две большие группы: витаминные и невитаминные. Коферменты витаминной природы – производные витаминов или химические модификации витаминов. 1 группа: тиаминовые – производные витамина В1. Сюда относят: - тиаминмонофосфат (ТМФ); - тиаминдифосфат (ТДФ) или тиаминпирофосфат (ТПФ) или кокарбоксилаза; - тиаминтрифосфат (ТТФ). ТПФ имеет наибольшее биологическое значение. Входит в состав декарбоксилазы кетокислот: ПВК, -кетоглутаровая кислота. Этот фермент катализирует отщепление СО. Кокарбоксилаза участвует в транскетолазной реакции из пентозофосфатного цикла. 2 группа: флавиновые коферменты, производные витамина В2. Сюда относят: - флавинмононуклеотид (ФМН); - флавинадениндинуклеотид (ФАД). Ребитол и изоалоксазин образуют витамин В2. Витамин В2 и остаток фосфорной к-ты образуют ФМН. ФМН в соединении с АМФ образуют ФАД. [рис. изоалоксазиновое кольцо соединено с ребитолом, ребитол с фосфорной к-той, а фосфорная к-та – с АМФ] 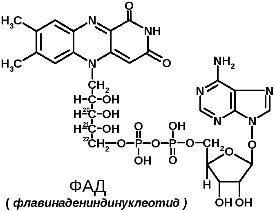 ФАД и ФМН являются коферментами дегидрогеназ. Эти ферменты катализируют отщепление от субстрата водорода, т.е. участвуют в реакциях окисления–восстановления. Например СДГ – сукцинатдегидрогеназа – катализирует превращение янтарной к-ты в фумаровую. Это ФАД-зависимый фермент. [рис. COOH-CH2-CH2-COOH (над стрелкой – СДГ, под – ФАД и ФАДН2) COOH-CH=CH-COOH]. Флавиновые ферменты (флавинзависимые ДГ) содержат ФАД, который в них является первоисточником протонов и электронов. В процессе хим. реакций ФАД превращается в ФАДН2. Рабочей частью ФАД является 2 кольцо изоалоксазина; в процессе хим. реакции идет присоединение двух атомов водорода к азотам и перегруппировка двойных связей в кольцах. 3 группа: пантотеновые коферменты, производные витамина В3 – пантотеновой кислоты. Входят в состав кофермента А, НS-КоА. Этот кофермент А является коферментом ацилтрансфераз, вместе с которой переносит различные группировки с одной молекулы на другую. 4 группа: никотинамидные, производные витамина РР - никотинамида: Представители: - никотинамидадениндинуклеотид (НАД); - никотинамидадениндинуклеотидфосфат (НАДФ). Коферменты НАД и НАДФ являются коферментами дегидрогеназ (НАДФ-зависимых ферментов), например малатДГ, изоцитратДГ, лактатДГ. Участвуют в процессах дегидрирования и в окислительно-восстановительных реакциях. При этом НАД присоединяет два протона и два электрона, и образуется НАДН2. 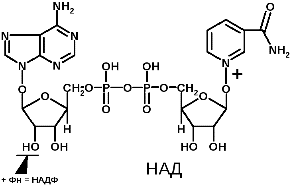 Рис. рабочей группы НАД и НАДФ: рисунок витамина РР, к которому присоединяется один атом Н и в результате происходит перегруппировка двойных связей. Рисуется новая конфигурация витамина РР + Н+] 5 группа: пиридоксиновые, производные витамина В6. [рис. пиридоксаля. Пиридоксаль+ фосфорная к-та= пиридоксальфосфат] - пиридоксин; - пиридоксаль; - пиридоксамин. Эти формы взаимопревращаются в процессе реакций. При взаимодействии пиридоксаля с фосфорной кислотой получается пиридоксальфосфат (ПФ). ПФ является коферментом аминотрансфераз, осуществляет перенос аминогруппы от АК на кетокислоту – реакция переаминирования. Также производные витамина В6 входят как коферменты в состав декарбоксилаз АК. Коферменты невитаминной природы – вещества, которые образуются в процессе метаболизма. 1) Нуклеотиды – УТФ, УДФ, ТТФ и т.д. УДФ-глюкоза вступает в синтез гликогена. УДФ-гиалуроновая к-та используется для обезвреживания различных веществ в трансверных реакциях (глюкоуронил трансфераза). 2) Производные порфирина (гем): каталаза, пероксидаза, цитохромы и т.д. 3) Пептиды. Глутатион – это трипептид (ГЛУ-ЦИС-ГЛИ), он участвует в о-в реакциях, является коферментом оксидоредуктаз (глутатионпероксидаза, глутатионредуктаза). 2GSH(над стрелкой 2Н) G-S-S-G. GSH является восстановленной формой глутатиона, а G-S-S-G – окисленной. 4) Ионы металлов, например Zn2+ входит в состав фермента АлДГ (алкогольдегидрогеназы), Cu2+ - амилазы, Mg2+ - АТФ-азы (например, миозиновой АТФ-азы). Могут участвовать в: -присоединении субстратного комплекса фермента; -в катализе; -стабилизация оптимальной конформации активного центра фермента; -стабилизация четвертичной структуры. | ||||||||||||||||||||||||||||||||||