Наследственная изменчивость. Комбинативная изменчивость
 Скачать 119.28 Kb. Скачать 119.28 Kb.
|

|
Наследственная изменчивость (неопределенная, индивидуальная, генотипическая) - форма изменчивости, вызванная изменениями генотипа организма, которые могут быть связаны с мутационной или комбинативной изменчивостью. Генотипическая изменчивость затрагивает генотип, а это значит, что генетические изменения затрагивают и половые клетки, которые передаются потомству. Поэтому и называется она - наследственная.  Комбинативная изменчивость Комбинативная изменчивость возникает в результате появления у потомков новых сочетаний генов (комбинаций). Эти комбинации возникают во время мейоза в результате кроссинговера - обмена участками между гомологичными хромосомами. В основе комбинативной изменчивости лежит три краеугольных момента: Случайная комбинация генов в ходе кроссинговера Независимое расхождение хромосом в мейозе Случайная встреча гамет при оплодотворении 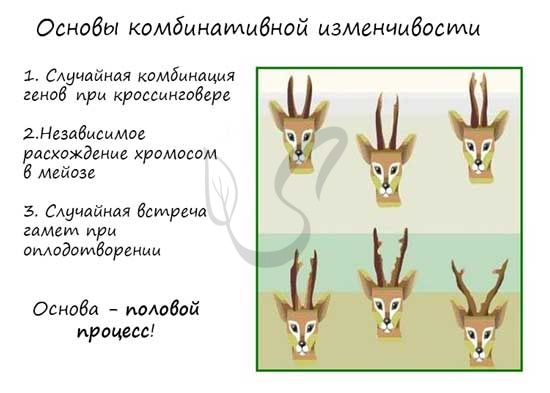 комбинативная изменчивость - это полная неопределенность: мы не знаем, какие комбинации возникнут между генами при кроссинговере, не знаем, какие хромосомы образуются и в какие гаметы они разойдутся, и, наконец, не знаем какие половые клетки (гаметы) встретятся при оплодотворении. То что мы отличаемся от своих родителей и наши дети отличаются от нас и есть результат неопределенностей Мутационная изменчивость Мутационная изменчивость связана с возникновением мутаций. Мутации (лат. mutatio - изменение) - внезапные, возникающие спонтанно или вызванные мутагенами наследуемые изменения генетического материала, приводящие к изменению тех или иных признаков организма. Мутации - резкие спонтанные изменения генотипа Стойкие, передаются потомкам через половые клетки (гаметы) Ненаправленные. Большинство мутаций - вредные (часть из них летальные), лишь очень небольшая часть носит полезный приспособительный характер, мутации также могут быть безразличными (нейтральными) для организма Носят индивидуальный характер. Среди мутаций различают следующие виды: Генные (точечные) Изменения при генных мутациях происходят в последовательности нуклеотидов молекулы ДНК. Может случаться такое, что один или несколько нуклеотидов выпадают из ДНК (делеция), вставляются новые нуклеотиды, удваиваются имеющиеся нуклеотиды (дупликация). Изменения ДНК ведут к тому, что в результате на рибосомах синтезируется белок с иной аминокислотной последовательностью. К примеру: изначально триплет ДНК "ТАЦ" кодировал аминокислоту "Мет", нуклеотид "Т" выпал из триплета произошла вставка нуклеотида "Г". В результате вместо аминокислоты "Мет" теперь синтезируется аминокислота Вал. Новые аминокислоты могут поменять свойства белка, так что признак, за который он отвечает, будет меняться. Хромосомные В результате хромосомных мутаций происходят структурные изменения хромосом (не следует путать с кроссинговером, который происходит в норме и подразумевает обмен участками между гомологичными хромосомами). Последствия хромосомных мутаций часто оказываются летальны. В результате таких мутаций может происходить утрата (делеция) участка хромосомы, его удвоение (дупликация), поворот на 180° (инверсия), перенос участка одной хромосомы на другую (транслокация), перенос участка внутри одной хромосомы (транспозиция). Геномные мутации Данный тип мутаций проявляется в изменении числа хромосом. Выделяют: Автополиплоидию - кратное увеличение числа наборов хромосом В результате таких мутаций количество хромосом увеличивается в кратное количество раз (2,3,4 и т.д.). В результате получаются организмы триплоиды, тетраплоиды и т.д. Иногда такие мутации вызывают искусственно, к примеру, в селекции растений. Известно, что у полиплоидов более крупные и сочные плоды. В селекции полиплоидию у растений вызывают добавлением специального химического вещества - колхицина, который блокирует образование нитей веретена деления. Вследствие этого хромосомы не расходятся и остаются в одной клетке - набор хромосом увеличивается в 2 раза. Аллополиплоидия (греч. állos — другой и polýploos — многократный) - объединение в организме хромосомных наборов от разных видов или родов Имеет значение в процессе видообразования. Примером данной мутации может послужить отдаленная гибридизация (аутбридинг) пшеницы и ржи. Их генотип состоит из гаплоидного набора пшеницы (n) и гаплоидного набора ржи (m). В результате такого скрещивания в 1875 году в Шотландии был получен первый искусственный стерильный гибрид - тритикале. Тритикале дает отличный урожай, в дальнейшем путем полиплоидии стерильность данного гибрида была преодолена. Также примером отдаленной гибридизации, соответственно и аллополиплоидии, является гибрид осла (самца) и лошади (самки) - мул. Это животное отличается большой выносливостью, но опять-тАнеуплоидия (греч. ἀν- — отрицательная приставка + εὖ — полностью + πλόος — кратный + εἶδος — вид Анеуплоидия - изменение кариотипа (совокупность признаков хромосом), при котором число хромосом в клетках не кратно гаплоидному набору (n). Таким образом, в результате анеуплоидии отсутствует одна (или несколько) хромосом, либо же хромосомы имеются в избытке ("лишние" хромосомы). В случае отсутствия в хромосомном наборе одной хромосомы говорят о моносомии, двух хромосом - нуллисомии. Если к паре хромосом добавляется одна лишняя, говорят о трисомии. Наследственные болезни, в том числе связанные с геномными мутациями: синдром Шерешевского-Тёрнера, Дауна - мы более детально обсудим в следующей статье, которая посвящена наследственным заболеваниям. аки бесплодное вследствие геномной мутации. Инбридинг (англ. in — в, внутри + breeding — разведение) - скрещивание близкородственных форм, в результате которого в ряду поколений увеличивается гомозиготность. С помощью инбридинга выводят чистые линии (AA, aa, BB, bb). Однако известно, что близкородственное скрещивание может приводить к проявлению рецессивных генов заболеваний и ослаблению потомства. Мутационный процесс. Изменения наследственного материала половых клеток в виде генных, хромосомных и геномных мутаций происходят постоянно. Особое место в этом процессе принадлежит генным мутациям, приводящим к возникновению серий аллелей и таким образом к разнообразию содержания биологической информации. Благодаря мутационному процессу поддерживается высокий уровень наследственного разнообразия природных популяций. Совокупность аллелей, возникающих в результате мутаций, составляет исходный элементарный эволюционный материал, который используется в процессе видообразования как основа действия других элементарных эволюционных факторов. Хотя отдельная мутация — событие редкое, общее число мутаций значительно. Большинство мутаций первоначально оказывает на фенотип особей неблагоприятное действие, однако, будучи рецессивными, мутантные аллели обычно присутствуют в генофондах популяций в гетерозиготных по соответствующему локусу генотипах. Благодаря этому достигается тройственный положительный результат: 1) исключается непосредственное отрицательное влияние мутантного аллеля на фенотипическое выражение признака, контролируемого данным геном; 2) путем сохранения аллелей, создается резерв наследственной изменчивости; 3) благодаря явлению гетерозиса (гибридной мощности) многие мутации, неблагоприятные по их прямому фенотипическому выражению, в гетерозиготном состоянии нередко повышают относительную жизнеспособность организмов. Хотя доля полезных мутаций мала, их абсолютное количество в пересчете на поколение или период существования вида может быть большим. Мутационный процесс, выполняя роль элементарного эволюционного фактора, происходит постоянно на протяжении всего периода существования жизни, а отдельные мутации возникают многократно у разных организмов. Популяционные волны. Популяционными волнами или «волнами жизни» (С. С. Четвериков) называют периодические или апериодические колебания численности организмов в природных популяциях. Это явление распространяется на все виды животных и растений, а также на микроорганизмы. Причины колебаний часто имеют экологическую природу. Так, размеры популяций «жертвы» (зайца) растут при ослаблении действия на них популяций «хищника» (рыси, лисицы, волка). Отмечаемое в этом случае увеличение кормовых ресурсов способствует росту численности хищников, что в свою очередь интенсифицирует истребление жертвы (рис. 107). Отдельные «вспышки» численности организмов некоторых видов, наблюдавшиеся в ряде регионов мира, были обусловлены деятельностью человека. В XIX—XXвеках это наблюдалось в популяциях кроликов в Австралии, домовых воробьев в Северной Америке, канадской элодеи в Евразии. Изменение генофондов популяций происходит как на подъеме, так и на спаде популяционной волны. При росте численности организмов наблюдается слияние ранее разобщенных популяций и объединение их генофондов. Так как популяции по своему генетическому составу уникальны, в результате такого слияния возникают новые генофонды с измененными по сравнению с исходными частотами аллелей. Рост количества организмов сопровождается расширением занимаемой территории. На гребне популяционной волны некоторые группы особей выселяются за пределы ареала вида, оказываются в нетипичных условиях существования и испытывают действие новых факторов естественного отбора. Повышение концентрации особей в связи с ростом их численности усиливает внутривидовую борьбу за существование. При спаде численности наблюдается распад крупных популяций. Возникающие малочисленные популяции характеризуются измененными генофондами. В условиях массовой гибели организмов редкие мутантные аллели могут быть генофондом вида потеряны. При сохранении редкого аллеля его концентрация в генофонде малочисленной популяции автоматически возрастает. На спаде «волны жизни» часть популяций, как правило, небольших по размерам, остается за пределами обычного ареала вида и, испытывая действие необычных условий жизни, вымирает. Иногда, благодаря благоприятному генетическому составу, такие популяции переживают период спада численности. Будучи изолированными от основной массы вида, существуя в необычной среде, они нередко служат родоначальниками новых видов. Действияпопуляционных волн на генофонды не являются направленными, поэтому они, так же как и мутационный процесс, подготавливают эволюционный материал к действию других элементарных эволюционных факторов. Изоляция. Только в результате прекращения панмиксии, т. е. благодаря изоляции, из одной исходной популяции или из групп могут сформироваться две или более генетически отличающиеся группы организмов, а в дальнейшем — новые подвиды и виды. Изоляция сама по себе не создает новых форм. Для их создания необходимы генетическая неоднородность и отбор, но изоляция способствует дивергенции. Выше уже говорилось, что, как правило, между видами отсутствует гибридизация, а следовательно, не происходит обмена генами. В этом смысле каждый вид представляет собой генетически закрытую систему. Представители различных популяций и подвидов, входящих в состав вида, легко скрещиваются между собой, обмениваются генами и, следовательно, являются генетически открытыми системами. Потенциально каждый подвид может дать начало новому виду, т. е. из генетически открытой системы перейти в генетически закрытую. В большинстве случаев, по-видимому, такому процессу способствует изоляция. Различают следующие основные формы изоляции: географическую, экологическую и генетическую. Географическая изоляция возникает в результате фрагментации ареала материнского вида. Она может быть следствием разграничения физическими барьерами (горными хребтами, водными пространствами и др.). Так возникли, например, эндемичные байкальские виды ресничных червей, ракообразных, рыб. Экологическая изоляция достигается тем, что две группы организмов, хотя и обитают в одной географической области, расселяются в различных экологических условиях или сроки размножения их не совпадают. Озерная и прудовая лягушки, нередко обитающие в одних водоемах, размножаются при различной температуре воды: первая приступает к икрометанию тогда, когда у второй оно закончено. Этим обеспечивается невозможность скрещивания между ними. Генетическая изоляция нередко обусловлена особенностями числа и формы хромосом, в силу чего у гибридов не может осуществляться мейоз (например, у мулов). Причинами изолирующего механизма становятся полиплоидия и хромосомные перестройки. В результате физиологической несовместимости тканей матери и гибридного эмбриона могут возникнуть препятствия для гибридизации у млекопитающих. В зависимости от характера изоляции различают две основные формы видообразования : аллопатрическое и симпатрическое. |