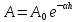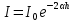КР1_Исин_8_вар. Контрольная работа По 1 модулю Мембранология и электрогенез. Механические колебания и волны. Агрегатные состояния вещества
 Скачать 75.68 Kb. Скачать 75.68 Kb.
|

1500 м/с). Однако высокая интенсивность и малая длина УЗ-волн порождают ряд специфических особенностей.Исин Куаныш Асанулы, 13-2 фип Контрольная работа По 1 модулю «Мембранология и электрогенез. Механические колебания и волны. Агрегатные состояния вещества» 1.Мембранный потенциал. Определение, величина. Уравнение Нернста. Потенциал Нернста, его природа. Стационарный мембранный потенциал. Каждая клетка превращает часть своей метаболической энергии в электростатическую энергию. Источником электрического поля клетки является плазматическая мембрана. Существует разность потенциалов между внутренней и внешней поверхностями плазматической мембраны. Эта разность потенциалов называется мембранным потенциалом. Разность потенциалов между внутренней и внешней средами клетки может измеряться непосредственно и довольно точно. Для этого используют микроэлектрод, представляющий собой стеклянную микропипетку с диаметром кончика до 1мкм, заполненную концентрированным раствором KCl. Микроэлектрод подключают к усилителю напряжения регистрирующего устройства. Можно измерять мембранный потенциал мышечных, нервных клеток или клеток других тканей. Другой электрод (референтный) установлен на поверхности ткани. Когда кончик микроэлектрода находится вне клетки, его потенциал по отношению к референтному электроду равен нулю. Если конец электрода погружают в клетку, прокалывая плазматическую мембрану, разность потенциалов резко становится отрицательной. На шкале измерительного устройства регистрируется разность потенциалов между внутренней и внешней средами клетки. Эта разность потенциалов называется трансмембранной, или мембранным потенциалом. Если клетка находится в состоянии покоя, её мембранный потенциал имеет отрицательное значение и устойчивую величину. Обычно его называют мембранным потенциалом покоя. Мембранный потенциал покоя клеток различных тканей составляет от-55 милливольт (мВ) до -100 мВ. При определенных физиологических условиях могут происходить изменения мембранного потенциала. Изменения его в положительном направлении называется деполяризацией плазматической мембраны. Смещение мембранного потенциала в отрицательном направлении называется гиперполяризацией. Электрические явления в плазматической мембране определяются распределением ионов между внутренней и внешней сторонами мембраны. Из химического анализа известно, что концентрация ионов внутриклеточной жидкости сильно отличается от концентрации ионов во внеклеточной жидкости. Термин "внеклеточная жидкость" имеет отношение ко всем жидкостям вне клеток (межклеточное вещество, кровь, лимфа и т.п.). Существуют значительные различия между концентрацией основных ионов внутри и вне клетки. Внеклеточная жидкость имеет высокую концентрацию ионов натрия и хлора. Внутриклеточная жидкость имеет высокую концентрацию калия и различных органических анионов (A-) (заряженные группы белков). Различие между концентрациями натрия и калия во внеклеточной и внутриклеточной жидкостях обусловлены деятельностью натрий-калиевого насоса, который выкачивает за один цикл 3 иона натрия из клетки и закачивает 2 иона калия в клетку против электрохимического градиента указанных ионов. Основная функция натрия-калия насоса - поддержание различия концентраций ионов натрия и калия по обе стороны плазматической мембраны. В состоянии покоя проницаемость плазматической мембраны для ионов калия значительно превышает проницаемость мембраны для ионов натрия. В нервных клетках соотношения проницаемости соответствующих ионов составляет 1:0,04. Этот факт дает возможность объяснять существование мембранного потенциала покоя. Ионы калия стремятся покинуть клетку из-за их высокой внутренней концентрации. При этом перемещения через мембрану внутриклеточных анионов из-за их больших размеров не происходит. Незначительное поступление ионов натрия внутрь клетки также не компенсирует выход ионов калия наружу, так как проницаемость мембраны в покое для ионов натрия мала. Следовательно, снаружи клетка приобретает дополнительно положительный заряд и внутри остаётся избыток отрицательного заряда. Диффузия калия через мембрану - процесс ограниченный. Ионы калия, проникающие через мембрану, создают электрическое поле, которое задерживает диффузию других ионов калия. По мере выхода из клетки калия электрическое поле нарастает и, в конечном итоге, напряжённость достигает такого значения, когда поток калия через мембрану прекращается. Состояние, при котором поток ионов по их концентрационному градиенту уравновешивается мембранным потенциалом, называется состоянием электрохимического равновесия ионов. Величина такого мембранного потенциала равновесия определяется уравнением Нернста (при этом считают, что мембрана проницаема только для одного вида ионов): 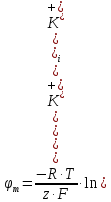 R - универсальная газовая постоянная, T - термодинамическая температура, z - электрический заряд иона, F - постоянная Фарадея, [K+]i и [K+]o - внутриклеточная и внеклеточная концентрации ионов калия соответственно. Вычисления, основанные на уравнении Нернста, указывают, что внутренняя и внешняя концентрация иона хлора также соответствует состоянию электрохимического равновесия, но концентрация натрия далека от равновесия с мембранным потенциалом мембраны. Уравнение Нернста показывает, что концентрационный градиент ионов калия определяет величину мембранного потенциала покоя только в первом приближении. Рассчитанные величины мембранного потенциала совпадают с экспериментально полученными только при высокой концентрации калия вне клетки. 2.Гидродинамика в одиночном сосуде. Уравнение Пуазейля. Гидравлическое сопротивление. Гемодинамика -- движение крови по сосудам, возникающее вследствие разности гидростатического давления в различных участках кровеносной системы (кровь движется из области высокого давления в область низкого). Одиночный сосуд рассматриваем как трубку кругового сечения, протяженную по сравнению со своими поперечными размерами. Под эластичностью стенок понимается возможность изменения сечения сосудов под действием давления. Пуайзель опытным путем установил, что средняя скорость ламинарного течения жидкости по неширокой горизонтальной круглой трубе постоянного сечения прямо пропорциональна разности давлений Р1 и Р2 при входе и выходи из трубы, квадрату радиуса трубы и обратно пропорциональна длине трубы l и вязкости n. 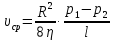 Гидравлическое сопротивление, сопротивление движению жидкостей (и газов) по трубам, каналам и т.д., обусловленное их вязкостью. Силой, создающей давление в сосудистой системе, является сердце. У человека среднего возраста при каждом сокращении сердца в сосудистую систему выталкивается 60--70 мл крови (систолический объем) или 4--5 л/мин (минутный объем). Движущей силой крови служат разность давлений, возникающая в начале и конце трубки. Почти во всех отделах сосудистой системы кровоток носит ламинарный характер -- кровь движется отдельными слоями параллельно оси сосуда. При этом слой, прилежащий к стенке сосуда, остается практически неподвижным, по этому слою скользит второй, а по нему, в свою очередь, третий и т. д. Форменные элементы крови составляют центральный, осевой поток, плазма движется ближе к стенке сосуда. Следовательно, чем меньше диаметр сосуда, тем ближе располагаются центральные слои к стенке и больше тормозится скорость их движения из--за вязкого взаимодействия со стенкой. В целом это означает, что в мелких сосудах скорость кровотока ниже, чем в крупных. В правильности этого положения легко убедиться, сопоставив скорости кровотока в разных участках сосудистого русла. В аорте она составляет 50--70 см/с, в артериях -- от 40 до 10, артериолах -- 10--0,1, капиллярах -- меньше 0.1, венулах -- меньше 0,3, венах -- 0,3--5,0, полой вене -- 5--20 см/с. Наряду с ламинарным в сосудистой системе существует турбулентное движение с характерным завихрением крови. Частицы крови перемещаются не только , параллельно оси сосуда, как при ламинарном кровотоке, но и перпендикулярно ей. Результатом такого сложного перемещения является значительное увеличение внутреннего трения жидкости. В этом случае объемная скорость тока крови будет уже не пропорциональной градиенту давления, а примерно равной квадратному корню из него. Турбулентное движение обычно возникает в местах разветвлений и сужений артерий, в участках крутых изгибов сосудов. Кровь представляет собой взвесь форменных элементов в коллоидно--солевом растворе, она обладает определенной вязкостью, не являющейся величиной постоянной. При протекании крови через капилляр, диаметр которого меньше 1 мм, вязкость уменьшается. Последующее уменьшение диаметра капилляра еще более уменьшает вязкость протекающей крови. Этот гемодинамический парадокс объясняется тем, что во время движения крови эритроциты сосредоточиваются в центре потока. Пристеночный же слой состоит из чистой плазмы с гораздо меньшей вязкостью, по которому легко скользят форменные элементы. В итоге улучшаются условия тока крови и происходит снижение перепадов давления, что, в общем, компенсирует увеличение вязкости крови и снижение скорости ее тока в мелких артериях. Переход от ламинарного движения крови к турбулентному сопровождается значительным ростом сопротивления течению крови. Соотношение между характером течения жидкости в жестких трубках и давлением обычно определяют по формуле Пуазейля. Используя эту формулу, можно вычислить сопротивление R току крови в зависимости от ее вязкости 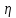 , длины l и радиуса r сосуда: , длины l и радиуса r сосуда: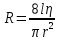 Сосудистую систему в целом можно представить в виде последовательно и параллельно соединенных трубок разной длины и диаметра. В случае последовательного соединения общее сопротивление составляет сумму сопротивлений отдельных сосудов: R = R1+ R2 + … + Rn. При параллельном соединении величину сопротивления вычисляют по другой формуле: 1/R = 1/R1 + 1/R2 + ... + + 1/Rn. Учитывая сложность геометрии сосудов целого организма, ее непостоянство, зависящее от открытия и закрытия шунтов, коллатералей, степени сокращения гладких мышц, эластичности стенок, изменения вязкости крови и других причин, в реальных условиях рассчитать величину сосудистого сопротивления трудно. Поэтому его принято определять как частное от деления кровяного давления Р на минутный объем крови Q: R = P/Q. Для всей сосудистой системы организма в целом эта формула применима лишь при том условии, если в конце системы, т. е. в полых венах вблизи места их впадения в сердце, давление будет близким к нулю. Соответственно при необходимости вычисления сопротивления отдельного участка сосудистой системы формула приобретает вид:  Значения P1 и P2 отражают давление в начале и конце определяемого участка. Основная кинетическая энергия, необходимая для движения крови, сообщается ей сердцем во время систолы. Одна часть этой энергии расходуется на проталкивание крови, другая -- превращается в потенциальную энергию растягиваемой во время систолы эластичной стенки аорты, крупных и средних артерий. Их свойства зависят от наличия эластических и коллагеновых волокон, растяжимость которых примерно в шесть раз выше, чем, например, резиновых нитей той же толщины. Во время диастолы энергия стенки аорты и сосудов переходит в кинетическую энергию движения крови. Кроме эластичности и растяжимости, т. е. пассивных свойств, сосуды обладают еще способностью активно реагировать на изменение в них кровяного давления. При повышении давления гладкие мышцы стенок сокращаются и диаметр сосуда уменьшается. Таким образом, пульсирующий ток крови, создаваемый функцией сердца, благодаря особенностям аорты и крупных сосудов выравнивается и становится относительно непрерывным. Основными показателями гемодинамики являются объемная скорость, скорость кругооборота крови, давление в разных областях сосудистой системы. Объемная скорость движения крови характеризует ее количество (в миллилитрах), протекающее через поперечное сечение сосуда за единицу времени (1 мин). Объемная скорость кровотока прямо пропорциональна перепаду давления в начале и конце сосуда и обратно пропорциональна его сопротивлению току крови. В организме отток крови от сердца соответствует ее притоку к нему. Это означает, что объем крови, протекающей за единицу времени через всю артериальную и всю венозную систему большого и малого круга кровообращения, одинаков. Линейная скорость движения крови (v) характеризует скорость перемещения ее частиц вдоль сосуда при ламинарном потоке. Она выражается в сантиметрах в секунду и определяется как отношение объемной скорости кровотока Q к площади поперечного сечения сосуда 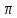 r2: r2: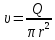 Полученная таким образом величина является сугубо средним показателем, так как, согласно законам ламинарного движения, скорость перемещения крови в центре сосуда является максимальной и падает в слоях, прилежащих к сосудистой стенке. Линейная скорость кровотока различна и в отдельных участках сосудистого русла по ходу сосудистого дерева. Она зависит от общей суммы площади просветов сосудов этого калибра в рассматриваемом участке. Наименьшим поперечным сечением характеризуется аорта, в связи с чем и скорость движения крови в ней самая большая - 50-70 см/с. Наибольшей суммарной площадью поперечного сечения обладают капилляры, у млекопитающих она приблизительно в 600-800 раз больше площади поперечного сечения аорты. Соответственно и скорость крови здесь около 0,05 см/с. В артериях она составляет 20--40 см/с, в артериолах - 0,5 см/с. В силу того, что при слиянии вен их суммарный просвет уменьшается, линейная скорость кровотока снова возрастает, достигая в полой вене 20 см/с (рис. 9.30). Кровь выталкивается отдельными порциями, поэтому кровоток в аорте и артериях пульсирует. При этом его линейная скорость возрастает в фазе систолы и снижается во время диастолы. В капиллярной сети в силу особенностей строения предшествующих ей артерий пульсовые толчки исчезают, и линейная скорость кровотока приобретает постоянный характер. 3.Ультразвук: способы получения (обратный пьезоэффект, магнитострикция) Ультразвук - упругие колебания и волны с частотами приблизительно от 20x103 Гц (20 кГц) и до 109 Гц (1 ГГц). Излучатели ультразвука Для получения ультразвука в медицине используют обратный пьезоэлектрический эффект. Он состоит в том, что пластинка, вырезанная определенным образом из кристалла кварца, под действием электрического поля деформируется в зависимости от направления поля. Если поместить такую пластинку между обкладками плоского конденсатора, на которые подаются переменное напряжение, то пластинка придет в вынужденные колебания. Эти колебания приобретают наибольшую амплитуду, когда частота изменений электрического напряжения совпадает с частотой собственных колебаний пластинки. Колебания пластинки передаются частицами окружающей среды, что и порождаются ультразвуковую волну.Некоторые материалы способны изменять свои размеры в магнитном поле. Это явление, получившее название магнитострикции, принципиально не отличается от обратного пьезоэлектрического эффекта. К числу подобных материалов относятся титанат бария и титанат-цирконат свинца. Преобразователи, в которых используются описанные выше явления, называют пьезоэлектрическими. Свойства ультразвука: 1) Поглощение ультразвука в веществе. По физической сущности УЗ не отличается от звука и представляет собой механическую волну. При ее распространении образуются чередующиеся участки сгущения и разряжения частиц среды. Скорость распространения УЗ и звука в средах одинаковы (в воздухе |