Гистологическое строение тимуса. Курсовая работа по гистологии, эмбриологии, цитологии Гистологическое строение тимуса Тема работы студент 22ЛЛ7 группы 1 курса Чернышов Руслан Мансурович Проверила
 Скачать 205.48 Kb. Скачать 205.48 Kb.
|

|
ФГБОУ ВО «Пензенский государственный университет» Медицинский институт Кафедра «Морфология» Курсовая работа по гистологии, эмбриологии, цитологии «Гистологическое строение тимуса» Тема работы Выполнил: студент 22ЛЛ7 группы 1 курса Чернышов Руслан Мансурович Проверила: Кандидат биологических наук, доцент Вишнякова Жанна Сергеевна Пенза 2023 СОДЕРЖАНИЕВВЕДЕНИЕ 3 ГЛАВА 1. ЭМБРИОГЕНЕЗ ТИМУСА 5 ГЛАВА 2. ГИСТОЛОГИЧЕСКОЕ СТРОЕНИЕ ТИМУСА 10 ГЛАВА 3. ФУНКЦИОНАЛЬНАЯ РОЛЬ ТИМУСА В ОРГАНИЗМЕ ЧЕЛОВЕКА 14 ГЛАВА 4. ИНВОЛЮЦИЯ ТИМУСА 18 4.1. Возрастная инволюция 18 4.2. Акцидентальная инволюция тимуса 19 ЗАКЛЮЧЕНИЕ 20 СПИСОК ЛИТЕРАТУРЫ 21 ВВЕДЕНИЕТимус — это центральный, или первичный, орган лимфоидной системы. Как известно, его основные функции заключаются в обеспечении созревания и дифференцировки Т-лимфоцитов, интеграции различных популяций Т-лимфоцитов и макрофагов для реализации иммунных ответов. Морфофункциональное состояние центрального органа во многом определяет активность вторичных структур иммуногенеза и выраженность защитных реакций всего организма. У всех млекопитающих животных тимус располагается в средостении, а у человека он находится в передней части верхнего средостения. К его вентральной поверхности прилежат тело грудины, грудино-подъязычная и грудино-щитовидная мышцы, париетальная плевра, а к дорсальной — перикард, трахея, дуга аорты, внутренние яремные и плечеголовные вены, возвратный гортанный нерв. Данный орган является предметом интенсивных исследований в области иммунологии, физиологии и гистологии на протяжении последних 60 лет. Однако пик интереса к организации и функционированию тимуса приходится на 1950-60-е гг., когда была установлена его роль в созревании и дифференцировке части лимфоидных клеток, получивших название Т-лимфоцитов. Классические исследования гистологии тимуса были проведены ещё раньше, в 1920-30-ее гг. и были связаны с работами немецкой гистологической школы. Общеизвестно, что тимус состоит из долей. Еще в XIX в. было отмечено, что количество долей в тимусе человека варьирует от одной, при слиянии правой и левой долей до пяти, что было подтверждено в XX в. Тем не менее считается, что основным вариантом строения тимуса человека является его двухдолевая организация. Цель курсовой работы: изучение гистологическое строения тимуса. Задачи курсовой работы: 1. Изучить эмбриогенез вилочковой железы; 2. Рассмотреть гистологическое строение тимуса; 3. Выделить основную функцию тимуса в иммунной системе. ГЛАВА 1. ЭМБРИОГЕНЕЗ ТИМУСАТимус в структурно-функциональном плане представляет собой сложную систему, выполняющую одновременно защитную и эндокринную функции. Являясь центральным органом иммуногенеза, он обеспечивает созревание Т-лимфоцитов, в частности появление у них анти-генраспознающих рецепторов, дифференцировку Т-лимфоцитов на субпопуляции и селекцию клонов Т-лимфоцитов, способных распознавать чужеродные пептиды в комплексе с аутологичными продуктами главного комплекса гистосовместимости. Как эндокринный орган, вилочковая железа синтезирует гормоны, основными из которых являются тимозин, тимоген, тимулин, тимопоэтин. На этапах эмбриогенеза в развитии иммунной системы и тимуса целом выделяют несколько критических периодов. Начало формирования тимуса приходится на 6-ю неделю эмбрионального развития, и до 8-й недели он имеет строение только эпителиального органа. На 7-й неделе гестации эпителиальные тяжи мигрируют за грудину, в анатомическую зону локализации тимуса. Закладка тимуса происходит на 4-й неделе эмбриогенеза в виде двух тяжей многослойного плоского эпителия, мигрирующих из III и частично IV пар жаберных карманов краниального отдела головной кишки. В пределах эпителиального пласта начинается дифференцировка клеток. Так, на периферии пласта эпителиальные клетки, преобразуясь в призматические, располагаются сплошным слоем на базальной мембране, а в центральной части тяжа они приобретают цитологические признаки ретикулоэпителия. Рост органа на 5-6-й неделях эмбриогенеза осуществляется за счет активного размножения расположенных на периферии эпителиальных клеток. Вокруг эпителиальных клеток формируется капсула, из которой кровеносные сосуды капиллярного типа врастают в эпителиальные тяжи. На этой стадии развития орган пока не заселён лимфоцитами. Появление Т-лимфоцитов в тимусе на 5-й неделе эмбриогенеза связывают с дистантным влиянием тимического фактора на стволовые клетки печени, которая на данном этапе онтогенеза является единственным органом кроветворения. Основные структуры тимуса формируются с 7-й по 12-ю неделю эмбрионального развития. Так, с 7-й по 8-ю неделю эпителиальная часть органа заселяется дифференцированными Т-лимфоцитами СР2. Дифференцировка лимфоцитов осуществляется в тесной взаимосвязи с ретикулоэпителием - одним из основных элементов клеточного микроокружения тимоцитов. К этом сроку в тимусе определяются первые кровеносные сосуды, через которые мигрируют пре-Т-лимфоциты. К 8-й неделе гестации клетки субкапсулярной зоны в виде широкого пласта мигрируют в окружающую мезенхиму и отграничивают участки мезенхимы с проходящими в ней кровеносными сосудами. При этом по всей поверхности сохраняется субкапсулярная зона. Результатом указанных выше процессов в развивающемся тимусе является формирование гематотимического барьера, который состоит из эндотелия, мезенхимы и прилегающих к ней эпителиальных клеток. Базальная мембрана с расположенными на ней эпителиальными клетками формирует барьерный комплекс, препятствующий проникновению в ретикулоэпителий различных макромолекул, в том числе и антигенов. Расщепление первоначально широких выростов ретикулоэпителия происходит на 10-й неделе. Несмотря на постоянную пролиферацию эпителиальных клеток, данная зона не увеличивается, поскольку орган непрерывно заселяется лимфоцитами, мигрирующими из центра дольки на периферию. Миграция лимфоцитов в формирующийся тимус связана с таким фактором, как ослабление межклеточных контактов за счет снижения пиронинофилии и содержания мукопроте-идов в эпителиальных клетках. Следовательно, клетки эпителия субкапсулярной зоны, обладающие большой пролиферативной потенцией, являются камбиальными и формируют основную зону роста долек тимуса на ранних этапах эмбриогенеза. Существование зон роста в субкапсулярной и центральной зонах тимуса обеспечивает быстрое увеличение массы органа на завершающем этапе эмбриогенеза. Если к 12-й неделе масса тимуса составляет 24,5 мг, то к 13-15-й она увеличивается в 5,3 раза и достигает 131 мг. Основу клеточного микроокружения лимфоцитов составляют эпителиальные клетки, которые к 8-9-й неделям гестации формируются в многочисленные соединения типа десмосом. Дифференцировку эпителиальных клеток на данном этапе развития определяют разные по размерам везикулы и тонофибриллы, расположенные в большом количества в цитоплазме. В указанные сроки начинается активная секреция клеток ретикулоэпителия, которая обеспечивает дифференцировку лимфоидных элементов и способствует миграции стволовых кроветворных клеток и пре-Т-лимфоцитов в орган. На 8-й неделе эмбриогенеза преобразуется мозговое вещество, в которое наряду с соединительной тканью и кровеносными сосудами врастают эпителиальные клетки субкапсулярной зоны, что свидетельствует о формировании в центре долек структурной основы гематотимического барьера и зоны роста эпителиальной стромы. С 12-й недели внутриутробного развития за счет миграции особых, так называемых «светлых эпителиальных клеток», в мозговом веществе образуются первые тимические тельца. Формирование телец Гассаля начинается с группировки эпителиальных клеток, кариопикноза и кари-орексиса их ядер, с последующим наслоением уплощенного эпителия и образованием структур слоистого типа. Изменения структурной организации, гистохимического состава и морфометрических параметров тимических телец отражают стадии их морфогенеза: прогрессивную (тип формирующихся и молодых телец), регрессивную, включающую фазу морфофункциональной зрелости (тип зрелых телец) и дегенерации (тип старых телец). Лимфоцитам тимуса, начиная с момента заселения органа, т. е. с 7,5 - 8-ми недель развития, свойственна морфологическая неоднородность. К10 - й неделе развития около 26% популяции составляют большие бластоподобные клетки диаметром 10-19 мкм неправильной или вытянутой формы с базофильной цитоплазмой и сетчатой структурой ядра. Другая часть клеток относится к средним лимфоцитам, имеющим округлую форму и слабо базофильную цитоплазму. К концу эмбриогенеза (12-я неделя) в тимусе содержится до 85% Т-лимфоцитов СO2 и до 1,3% В-лимфоцитов. На 13-й неделе развития появляются малые лимфоциты, которые на последующих этапах фетогенеза становятся основными клеточными элементами и обеспечивают формирование Т-клеточной системы иммунитета. На 9-й неделе развития в междольковых прослойках соединительной ткани тимуса появляются первые гранулоциты, а на 12-й - макрофаги и тканевые базофилы. В перечисленных стромальных клетках синтезируются различные гуморальные факторы, способствующие дифференцировке тимоцитов и пролиферации лимфоидных элементов. Все этапы роста и дифференцировки клеток тимуса регулируются огромным количеством разнообразных по функциональной направленности биологически активных веществ. Последние подразделяются на регуляторные - тимозин а-1,3, 7, 11, тимозины. Таким образом, вторым критическим периодом в морфогенезе тимуса является стадия 7-12-й недель гестации, характеризующаяся образованием долек с дифференцировкой на корковое и мозговое вещество, формированием телец Гассаля, зон роста на периферии и в центре долек, а также дифференцировкой лимфоцитов и последующей миграцией их в периферические органы иммунной системы. На протяжении последующих недель развития, 14-17-я недели, центрального органа иммунной системы завершается формирование четырех структурно-функциональных зон: субкапсулярной, внутренней кортикальной, медуллярной и внутридольковой периваскулярной. В субкапсулярной зоне осуществляется пролиферация и начальная дифференцировка предшественников Т-лимфоцитов. Благодаря прямому контакту эпителиальных клеток (клетки-«няньки») с лимфоцитами и влиянию на них местнодействующих тимических гормонов продолжается дальнейшая дифференцировка Т-лимфоцитов. В структурах внутренней кортикальной зоны происходит созревание и селекция аутотолерантных Т-лимфоцитов. Медуллярная зона содержит зрелые Т-лимфоциты. Четвертой структурно-функциональной зоной тимуса являются внутридольковые периваскулярные пространства, представляющие собой не что иное, как транспортные пути для миграции лимфоцитов. По завершении формообразования составных компонентов тимуса дальнейший рост центрального органа иммуногенеза осуществляется неравномерно с пиками в 12-15 и затем в 16-21 неделю. Темпы роста тимуса, максимальные на 19-24-й неделях, постепенно и последовательно снижаются к 30-й неделе. Осуществляющаяся до 22-й недели гестации внутридольковая дифференцировка тимуса далека от завершения, поэтому мозговое вещество в органе преобладает над корковым. Относительное увеличение объёма мозгового вещества в тимусе обусловлено продолжающейся дифференцировкой ретикулоэпителиальных, стромальных клеток и значительным увеличением количества и размеров телец Гассаля, которые располагаются в указанные сроки как в центре, так и на периферии органа. ГЛАВА 2. ГИСТОЛОГИЧЕСКОЕ СТРОЕНИЕ ТИМУСА Снаружи тимус покрыт соединительнотканной капсулой. От нее внутрь отходят перегородки, разделяющие его на дольки. В каждой дольке различают корковое и мозговое вещество. Долька органа включает эпителиальную ткань, состоящую из отростчатых клеток - ретикулярных эпителиоцитов, а также клеток моноцитоидного происхождения. Для всех ретикулярных эпителиоцитов характерно наличие десмосом, тоно-филаментов и белков кератинов, продуктов главного комплекса гистосовме-стимости I и II классов в составе плазматической мембраны. Ретикулярные эпителиоцитыв зависимости от локализации отличаются формой и размерами, тинкториальными признаками, электронной плотностью гиалоплазмы, содержанием органелл и включений. Описаны секреторные клетки коры и мозгового вещества, несекреторные (опорные) и клетки эпителиальных слоистых телец - телец Гассаля (гассалевы тельца), периваскулярные. Секреторные клетки содержат вакуоли или секреторные включения. При помощи моноклональных антител в них обнаружены гормоноподобные факторы: α-тимозин, тимусный сывороточный фактор, тимопоэтины. Эпителиальные клетки в подкапсулярной зоне и в части коркового вещества имеют глубокие инвагинации, в которых расположены, как в колыбели, лимфоциты. Прослойки цитоплазмы этих эпителиоцитов - тимусных питающих клеток, или «нянек», между лимфоцитами могут быть очень тонкими и протяженными. Обычно такие клетки содержат 10-20 лимфоцитов и более. Корковое вещество (cortex) - периферическая часть долек, содержит Т-лимфоциты, которые густо заполняют просветы сетевидной эпителиальной основы. В подкапсулярной зоне коркового вещества находятся крупные лимфоидные клетки - лимфобласты - предшественники Т-лимфоцитов, мигрировавшие сюда из красного костного мозга. Они под влиянием тимозина, выделяемого ретикулярными эпителиоцитами, пролиферируют. Новые генерации лимфоцитов появляются в тимусе каждые 6-9 ч. Созревая, Т-лимфоциты перемещаются в направлении мозгового вещества и покидают тимус через венулы кортикомедуллярной зоны. Полагают, что Т-лимфоциты коркового вещества мигрируют в кровоток, не входя в мозговое вещество. Эти лимфоциты отличаются по составу рецепторов от Т-лимфоцитов мозгового вещества. С током крови они попадают в периферические органы лимфоцитопоэза - лимфатические узлы и селезенку, где созревают в субклассы: антиген-реактивные киллеры, хелперы, супрессоры. Однако не все образующиеся в тимусе лимфоциты выходят в циркуляторное русло, а лишь около 2-5 %, которые прошли «обучение» и приобрели специфические циторецепторы чужеродных антигенов. Лимфоциты, имеющие циторецепторы собственных антигенов, как правило, погибают в тимусе, что служит проявлением отбора иммунокомпетентных клеток. При попадании этих Т-лимфоцитов в кровоток развивается аутоиммунная реакция. Клетки коркового вещества отграничены от крови гематотимусным барьером, предохраняющим дифференцирующиеся лимфоциты коркового вещества от избытка антигенов. В его состав входятэндотелиальные клетки капилляров с базальной мембраной, перикапиллярное пространство с единичными лимфоцитами, макрофагами, фибробластами и межклеточным веществом, а также специализированные ретикулярные эпителиоциты с их базальной мембраной. Барьер обладает избирательной проницаемостью по отношению к антигену. При нарушении барьера среди клеточных элементов коркового вещества обнаруживаются единичные плазматические клетки, зернистые лейкоциты и тучные клетки. Иногда в корковом веществе появляются очаги экстрамедуллярного миелопоэза. 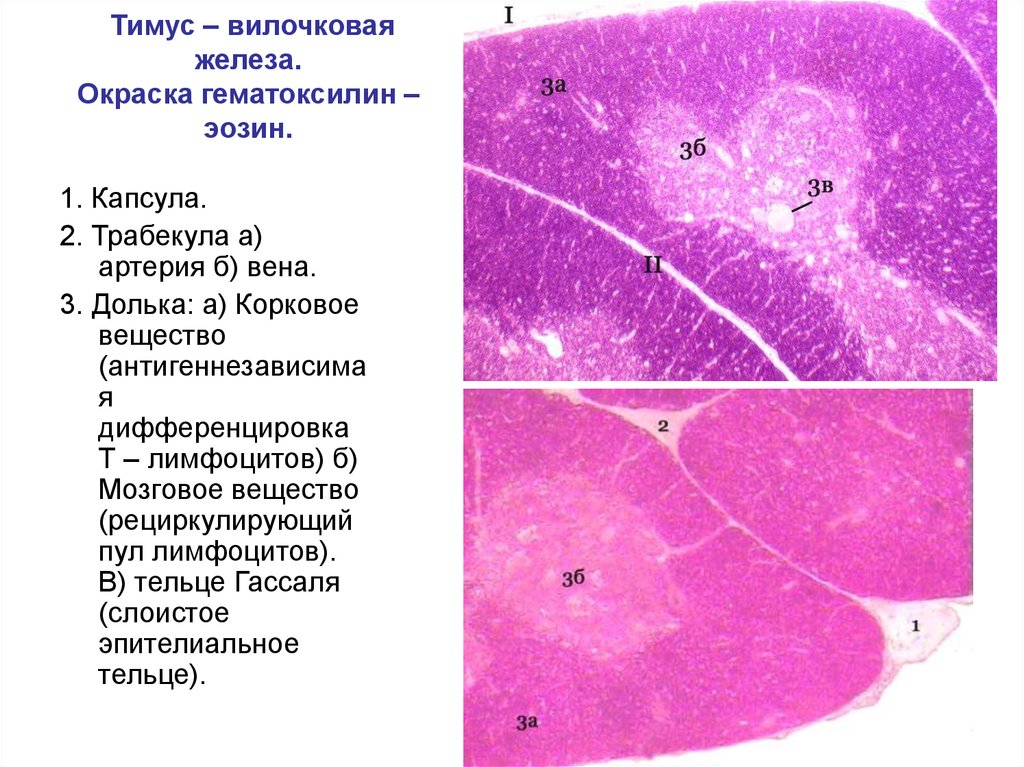 Рис. №1 «Вилочковая железа (тимус) 1. – капсула, 2 – трабекура, 3а – корковое вещество, 3б – мозговое вещество, 3в – тельца Гассаля Мозговое вещество (medulla) дольки на гистологических препаратах имеет более светлую окраску, так как по сравнению с корковым веществом содержит меньшее количество лимфоцитов. Лимфоциты этой зоны представляют собой рециркулирующий пул Т-лимфоцитов и могут поступать в кровь и выходить из кровотока через посткапиллярные венулы. Количество митотически делящихся клеток в мозговом веществе примерно в 15 раз меньше, чем в корковом. Особенностью ультрамикроскопического строения ретикулярных эпителиоцитов является наличие в их цитоплазме гроздевидных вакуолей и внутриклеточных канальцев, поверхность которых образует микровыросты. В средней части мозгового вещества расположены слоистые эпителиальные тельца (corpusculum thymicum).Они образованы концентрически наслоенными ретикулярными эпителиоцитами, цитоплазма которых содержит крупные вакуоли, гранулы кератина и пучки фибрилл. Они хорошо развиты у человека, собаки, морской свинки и слабо развиты у мышей и крыс. Количество этих телец у человека увеличивается к периоду половой зрелости, затем уменьшается. Функция телец не установлена. Внутри органа артерии ветвятся на междольковые и вну-тридольковые, которые образуют дуговые ветви. От них почти под прямым углом отходят кровеносные капилляры, образующие густую сеть, особенно в корковом веществе. Капилляры коркового вещества окружены непрерывной базальной мембраной и слоем эпителиальных клеток, отграничивающим перикапиллярное пространство. В перикапиллярном пространстве, заполненном тканевой жидкостью, встречаются лимфоциты и макрофаги. Большая часть корковых капилляров переходит непосредственно в подкап-сулярные венулы. Меньшая часть идет в мозговое вещество и на границе с корковым веществом переходит в посткапиллярные венулы, отличающиеся от капсулярных венул высоким призматическим эндотелием. Через этот эндотелий могут рециркулировать (уходить из тимуса и вновь возвращаться) лимфоциты. Барьера вокруг капилляров в мозговом веществе нет. Лимфатическая система представлена глубокой и поверхностной (кап-сулярной и подкапсулярной) выносящей сетью капилляров. Глубокая капиллярная сеть особенно богата в корковом веществе, а в мозговом веществе капилляры обнаружены вокруг эпителиальных слоистых телец. Лимфатические капилляры собираются в сосуды междольковых перегородок, идущие вдоль кровеносных сосудов. ГЛАВА 3. ФУНКЦИОНАЛЬНАЯ РОЛЬ ТИМУСА В ОРГАНИЗМЕ ЧЕЛОВЕКА Тимус – один из центральных органов иммунной системы. Из-за большого количества апоптотических лимфоцитов первоначально ему предписывалась функция «кладбища» лимфоцитов. Роль тимуса в иммунной системе была продемонстрирована лишь в 1961 г. Жаком Миллером. Он показал, что в тимусе Т-клетки-прекурсоры, мигрировавшие из костного мозга, подвергаются селекции и созреванию с последующей миграцией в Т-клеточнозависимые области периферических органов лимфоидной системы – селезенки, лимфатических узлов, Пейеровых бляшек и нёбных миндалин. Процесс дифференциации включает экспрессию различных мембранных маркеров и реорганизацию Т-клеточных рецепторов. Следует отметить, что, хотя активность пролиферации и дифференцировки тимоцитов снижается с возрастом, ВЖ в течение всей жизни обеспечивает созревание и миграцию Т-лимфоцитов к периферическим органам иммунной системы. Различные индуктивные, гормональные и пролиферативные сигналы от эпителиальных клеток способствуют созреванию тимоцитов. В процессе созревания взаимодействие Т-клеток и эпителиоцитов происходит посредством антигенов главного комплекса гистосовместимости. Интратимическое созревание αβ-Т-клеток основывается на функциональном балансе позитивной и негативной селекции. Для обеспечения эффективного иммунного ответа, помимо образования функционально разнообразных клонов T-клеток, необходим механизм ограничения активации случайно генерируемых аутореактивных Т-клеток. Эта функция тимуса важна на протяжении всей жизни человека и обеспечивается различными эпителиальными клетками. Кортикальные тимические эпителиальные клетки (cTEC) отвечают за позитивную селекцию. Напротив, медуллярные эпителиальные клетки тимуса (mTEC) ответственны за внутриорганную толерантность к Т-клеткам и негативную селекцию аутоагрессивных клонов, а также созревание регуляторных Т-клеток. Нарушение вышеописанных механизмов может привести к аутоиммунным заболеваниям. Так, аутоиммунная болезнь человека (APECED) связана с аберрантным развитием и нарушением функции медуллярного тимуса в виде дефекта зрелых mTEC и мутаций в аутоиммунном регуляторе. О роли тимуса в поддержании гомеостаза организма, помимо иммунной функции, стало известно после получения популяции тимэктомированных подопытных мышей. Помимо иммунодефицита, они характеризовались полным отсутствием оволосения, значительным уменьшением массы и длины тела, гипотрофическими изменениями в органах и тканях, нарушением строения костей скелета. Следует отметить, что удаление тимуса у взрослых мышей не приводит к формированию иммунодефицита, а ассоциируется лишь со снижением популяции лимфоцитов в крови, лимфомой грудного протока, лимфатических узлов и селезенки. На основании этих данных впервые предположили, что в раннем возрасте тимус необходим для полного и нормального развития некоторых иммунологических реакций. Тимус у взрослых, по-видимому, необходим для коррекции специфических иммунологических дефектов. Перекрестные взаимодействия между нейроэндокринной и иммунной системами в настоящее время активно изучаются. В этих системах используются гомологичные лиганды и рецепторы для установления физиологических интра- и интерсистемных взаимодействий, обеспечивающих поддержание гомеостаза. Физиологические реакции тимуса, как части единой нейроэндокринноиммунной системы, модулируются путем воздействия стероидных и полипептидных гормонов в различных биологических циклах. Эти эффекты реализуются посредством рецепторов к гормонам, идентифицированных на клетках тимуса. На сегодняшний день очевидны множественные взаимодействия между железами внутренней секреции и тимусом, однако полностью физиологическое значение этих связей не раскрыто. Так, щитовидная железа и гипофиз стимулируют секрецию тимулина. Напротив, надпочечники и половые железы находятся в антагонизме с тимусом. Предполагается связь снижения тимической секреции и уровня половых гормонов, которые начинают поступать в кровоток в период полового созревания. Тимус продуцирует ряд нейроэндокринноиммунных сигнальных молекул, к которым относят собственно гормоны тимуса, биогенные амины и пептидные гормоны APUD-клеток, лимфоцитов и клеток микроокружения. Также в тимусе синтезируются различные лимфокины, нейропептиды (нейротензин, субстанция Р, ВИП, холецистокинин, соматостатин, окситоцин, вазопрессин, нейротензины, метэнкефалин, АКТГ, предсердный натрийуретический пептид). Цитокины и тимические гормоны действуют аутокринным и паракринным путем, влияя на дифференцировку Т-лимфоцитов и гемопоэтические клетки. К основным гормонам тимуса относятся тимозин, тимопоэтин и тимулин. Тимопоэтин в первую очередь контролирует дифференцировку Т-лимфоцитов. Он также усиливает экспрессию проопиомеланокортина, секрецию продуктов его процессинга – АКТГ, -эндорфина и -липотропина, а также СТГ и кортизола. Тимозин непосредственно воздействует на клетки, обладающие иммунной реактивностью. Механизм действия тимозина основан на стимуляции лимфопоэза. Факт отсутствия иммунной реактивности лимфоцитов непосредственно в тимусе может быть объяснен тем, что клетки покидают тимус до стадии полного созревания. Функционально полноценными клетки становятся вне органа также под воздействием тимозина. В экспериментах при введении в желудочки мозга тимозин усиливает выработку гипоталамических рилизинг-факторов. На синаптических мембранах нейронов обнаружены рецепторы 1-тимозина. Тимулин усиливает выработку СТГ и пролактина. In vitro он усиливает секрецию прогестерона и эстрадиола гранулезными клетками, проявляя активность гонадотропин-рилизинг фактора. ГЛАВА 4. ИНВОЛЮЦИЯ ТИМУСА 4.1. Возрастная инволюция Тимус подвергается возрастной инволюции. Сроки этого процесса индивидуальны и в некоторой степени зависят от уровня половых гормонов, секретируемых в кровь во время периода полового созревания. Развитие тимуса происходит на 6–16-й неделях беременности, в перинатальном периоде орган достигает функциональной зрелости. Размеры железы достигают максимума (30–40 г) в раннем возрасте, затем происходит постепенное замещение функционально активной ткани тимуса белой жировой тканью, наиболее активно протекающее в пубертате. Размеры тимуса у взрослых значительно меньше по сравнению с детьми, тогда как количество нефункционирующей ткани намного выше. Кроме того, тимус теряет организационную архитектонику. Поэтому степень возрастной инволюции нельзя определить на основании массы или относительной плотности тимуса, которые могут сохраниться за счет жировой и соединительной ткани, а атрофические процессы при этом затрагивают только специфическую паренхиму железы. Существует несколько возможных объяснений возрастной инволюции органа, таких как дефицит Т-клеток-предшественников, мигрирующих из костного мозга, влияние циркулирующих гормонов и цитокинов. Однако ни одна из теорий точно не объясняет, почему функция тимуса уменьшается с возрастом. Недифференцированные Т-клетки также необходимы во взрослой жизни – они остаются в коммуникации и взаимозависимы с эпителиальными клетками тимуса во все периоды жизни человека. Однако известно о качественном снижении производства клеток-прекурсоров с возрастом. Нехватка наивных Т-клеток ассоциирована с опасностью тяжелого течения инфекций у пожилых. Содержание гормонов тимуса в циркуляции изменяется с возрастом. Максимальный уровень выработки гормонов тимуса наблюдается в раннем возрасте – до начала полового созревания. Затем секреторная активность клеток тимуса и концентрация гормонов в сыворотке крови постепенно снижаются. Это снижение также играет роль в возрастном влиянии тимуса на иммунную систему. 4.2. Акцидентальная инволюция тимуса Тимус чувствителен к любому виду стресса – инфекциям, новообразованиям, хирургическим вмешательствам, химиотерапии, ожогам. Реакция тимуса на стресс универсальна и проявляется быстрым уменьшением его размеров (до 40% от первоначальной массы) с последующим восстановлением или увеличением (более 50%, «rebound hyperplasia»). Временная, быстрая, или акцидентальная, инволюция может наступить в связи с воздействием на организм различных чрезвычайно сильных раздражителей (травма, интоксикация, инфекция, голодание и др.). При стресс-реакции происходят выброс Т-лимфоцитов в кровь и массовая гибель лимфоцитов в самом органе, особенно в корковом веществе. В связи с этим становится менее заметной граница коркового и мозгового вещества. Кроме лимфоцитолиза, наблюдается фагоцитоз макрофагами внешне не измененных лимфоцитов. Биологический смысл лимфоцитолиза окончательно не установлен. Вероятно, гибель лимфоцитов является выражением селекции Т-лимфоцитов. Одновременно с гибелью лимфоцитов происходит разрастание эпителиальной части органа. Эпителиоциты набухают, в цитоплазме появляются секретоподобные капли, дающие положительную реакцию на гликопротеины. В некоторых случаях они скапливаются между клетками, образуя подобие фолликулов. Тимус вовлекается в стресс-реакции вместе с надпочечниками. Увеличение в организме количества гормонов коры надпочечника, в первую очередь глюкокортикоидов, вызывает очень быструю и сильную акцидентальную инволюцию тимуса. ЗАКЛЮЧЕНИЕ Таким образом, тимус является важнейшим органом для организма, так как он является центральным органом иммунной системы, системы, которая защищает организм от чужеродных веществ и организмов, от собственных но переродившихся клеток. Именно тимус или вилочковая железа играет главную роль в этих процессах и, малейшее отклонение в ней неизбежно приведёт к нарушению иммунитета. В заключение отмечу, что в области строения, развития и функции вилочковой железы наибольшие достижения достигнуты только в последние десятилетия и многие вопросы остаются до конца не изученными. СПИСОК ЛИТЕРАТУРЫ 1. Гистология: Учебник/Ю. И. Афанасьев, Н. А. Юрина, Е. Ф. Котовский и др.; Под ред. Ю. И. Афанасьева, Н. А. Юриной. - 5-е изд., переаб. и доп. - М.: Медицина, 2002. - 744 с. 2. Гистология, эмбриология, цитология: учебник для вузов / Под ред. Э.Г.Улумбекова, Ю.А.Челышева - 3-е изд., - 2009. – 960 с. 3. Гистология, цитология и эмбриология. Атлас: учебное пособие. Быков В.Л., Юшканцева С.И. 2013. – 292 с. 4. Атлас по гистологии, цитологии и эмбриологии: учебное пособие /Кузнецов С.Л., Мушкамбаров Н.Н., Горячкина В.Л.- М.: МИА, 2002. – 172 с. 5. Цитология и общая гистология. Функциональная морфология клеток и тканей человека / В.Л. Быков // – СПб.: Сотис., - 2007. – 520 с. 6. Атлас по гистологии: Учебное пособие/Ред. А.С. Пуликов, Т.Г. Брюховец; Колл. авт. Красноярская мед. академия. – Красноярск: КрасГМА, 2004. - 128с 7. Курс возрастной гистологии:Учебное пособие/Ред. А.С. Пуликов; Колл. авт. Красноярская мед. академия. – Красноярск: ИПЦ «Версо», 2006. – 130с. 8. Гистология, цитология и эмбриология / С.И.Юшканцева, В.Л. Быков. Краткий атлас: Учебное пособие. 2-е изд., перераб. и доп. – СПб: Издательство: «П-2», 2007. – 120с. 9. Lele SM, Lele MS, Anderson VM. The thymus in infancy and childhood: embryologic, anatomic, and pathologic considerations. Chest. Surg. Clin. N. Am. 2001; 11: 233–253. 10. Zygmunt Zdrojewicz, Ewelina Pachura, Paulina Pachura. The Thymus: A Forgotten, But Very Important Organ. Adv. Clin. Exp. Med. 2016; 25 (2): 369–375. |