Занятие 21 обмен и функции углеводов метаболизм молочной кислоты, цикл Кори
 Скачать 343.3 Kb. Скачать 343.3 Kb.
|

|
БИОЛОГИЧЕСКАЯ ХИМИЯ Занятие 21 ОБМЕН И ФУНКЦИИ УГЛЕВОДОВ Метаболизм молочной кислоты, цикл Кори. Цикл Кори — совокупность биохимических ферментативных процессов транспорта лактата из мышц в печень, и дальнейшего синтеза глюкозы из лактата, катализируемое ферментами глюконеогенеза. При интенсивной мышечной работе, а также в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах или мышцах) глюкоза вступает на путь анаэробного гликолиза с образованием лактата. Лактат не может далее окисляться, он накапливается (при его накоплении в мышцах раздражаются чувствительные нервные окончания, что вызывает характерное жжение в мышцах). С током крови лактат поступает в печень. Печень является основным местом скопления ферментов глюконеогенеза (синтез глюкозы из неуглеводных соединений), и лактат идет на синтез глюкозы. Реакция превращения лактата в пируват катализируется лактатдегидрогеназой, далее пируват подвергается окислительному декарбоксилированию или может подвергаться брожению. Цикл Кори получил название по первооткрывателю — его открыла чешская ученая, лауреат Нобелевской премии Тереза Кори. 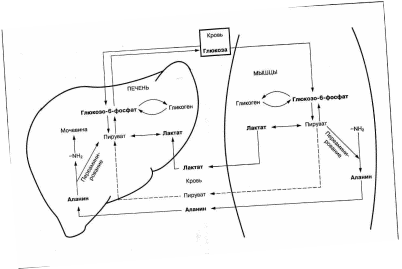 Глюконеогенез, метаболические предшественники глюкозы, схема глюконеогенеза, биологическая роль, регуляция. Глюконеогенез – это синтез глюкозы из неуглеводных предшественников, главным образом, из глицерина, молочной кислоты, гликогенных аминокислот, а также из ряда других молекул, способных превращаться в пируват, оксалоацетат (ЩУК) или промежуточные метаболиты цикла Кребса. Наиболее активно глюконеогенез протекает в печени, корковом веществе почек и слизистой оболочке тонкого кишечника. Центральным органом глюконеогенеза является печень – только она может продуцировать глюкозу «на экспорт», т.е. выделять образованную глюкозу в кровь. Большинство реакций глюконеогенеза представляет собой обратную последовательность реакций анаэробного гликолиза, за исключением трех необратимых реакций, которые в глюконеогенезе катализируются другими ферментами и называются ключевыми (основными) реакциями глюконеогенеза табл. 3. Эти уникальные для глюконеогенеза ферменты позволяют обойти необратимые реакции гликолиза. Таблица 3. Необратимые (ключевые) реакции гликолиза и глюконеогенеза
Большинство реакций глюконеогенеза протекает в цитоплазме. И только реакция превращения пирувата в ЩУК идет в митохондриях. Пируват, образованный в цитоплазме, транспортируется в митохондрию при помощи переносчика. В митохондрии пируват превращается в оксалоацетат ферментом пируваткарбоксилазой. Мембрана митохондрий не проницаема для оксалоацетата, поэтому он сначала превращается в малат под действием митохондриальной малатдегидрогеназы. Малат легко проходит через мембрану митохондрий и проникает в цитоплазму, где превращается в оксалоацетат под действием цитоплазмитической малатдегидрогеназы. Затем оксалоацетат превращается в фосфоенолпируват и далее – по пути глюконеогенеза (рис. 24). 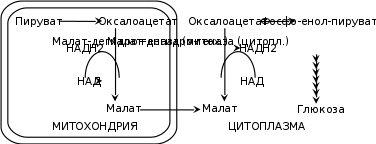 Рис. 24. Схема образования фосфоенолпирувата из пирувата в глюконеогенезе. Схема глюконеогенеза представлена на рис. 23. 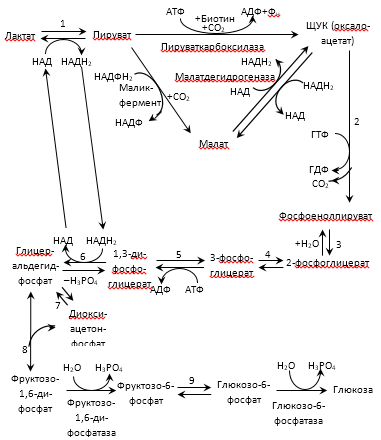 Рис. 23 Схема Глюконеогенеза  Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты цитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных. Например, окса-лоацетат, образующийся из аспарагиновой кислоты, является промежуточным продуктом как цитратногр цикла, так и глюконеогенеза. Биологическая роль глюконеогенеза 1. Во время голодания глюконеогенез поддерживает уровень глюкозы крови. В течение первых суток голодания концентрация глюкозы крови поддерживается на нормальном уровне благодаря распаду гликогена в печени. Но даже еще до полного истощения запасов гликогена в печени происходит активация глюконеогенеза. В среднем, начиная со 2-х суток голодания глюконеогенез становится единственным источником глюкозы крови и обеспечивает углеводное питание тканей (головной мозг), для которых глюкоза является единственным энергетическим субстратом. 2. При сахарном диабете активация глюконеогенеза является главной причиной развития гипергликемии. Среди причин развития гипергликемии при сахарном диабете доля различных механизмов может быть представлена следующим образом: 50 % – активация глюконеогенеза в печени; 20 % – активация глюконеогенеза в почках (при сахарном диабете корковое вещество почек начинает продуцировать глюкозу «на экспорт»); 30 % – другие механизмы, такие как - снижение транспорта глюкозы в клетку (уменьшение проницаемости клеточных мембран); - активация распада гликогена; - уменьшение синтеза гликогена; - торможение гликолиза. Регуляция глюконеогенеза Глюконеогенез и гликолиз регулируются противоположным образом, поэтому если один из путей активен, другой будет ингибирован. Инсулин тормозит глюконеогенез путем ингибирования ключевых ферментов: глюкозо-6-фосфатазы, фруктозо-1,6-дифосфатазы и фосфоенолпируваткарбоксикиназы. Глюкагон и глюкокортикоиды (кортизол) активируют глюконеогенез (ключевые ферменты. Аллостерическим активатором пируваткарбоксилазы является ацетил-КоА. Основные реакции глюконеогенеза. Роль биотина. 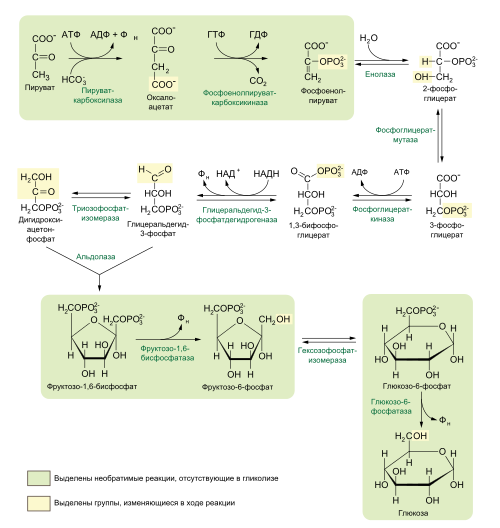 Роль биотина Фермент пируваткарбоксилаза содержит витамин Н, или биотин. Роль этого витамина заключается в том, что биотин связывает молекулу СО2 и делает ее активной, благодаря чему CO2 используется для наращивания углеродного скелета, в частности, пирувата при превращении его в оксалоацетат. Пентозофосфатный путь (ПФП), окислительные и неокислительные реакции, биологическая роль. В среднем, около 10 % глюкозы метаболизирует по этому пути. ПФП происходит в цитоплазме практически во всех клетках организма. Наиболее активно он протекает в эритроцитах, печени, жировой ткани, эндокринных железах (коре надпочечников, яичках, щитовидной железе), лактирующей молочной железе, эмбриональных тканях. В печени и эритроцитах до 30 % глюкозы метаболизирует по пентозофосфатному пути. Все реакции ПФП можно разделить на окислительные и неокислительные. На стадии окислительных (необратимых) реакций происходит окисление глюкозо-6-фосфата с образованием 2 молекул НАДФН2 и одной молекулы пентозофосфата – рибулозо-5-фосфата, при этом выделяется 1 молекула СО2. В неокислительных обратимых реакциях пентозофосфаты превращаются в промежуточные продукты гликолиза (такие как глицеральдегидфосфат и фруктозо-6-фосфат). Общий план расположения окислительных и неокислительных реакций ПФП представлен на рис 26.  Рис. 26. Относительное расположение окислительных и неокислительных реакций пентозофосфатного пути. Схема ПФП представлена на рис. 27, реакции – на рис. 28. 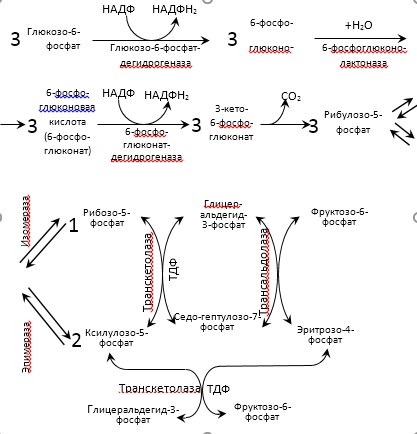 Рис. 27 Схема ПФП 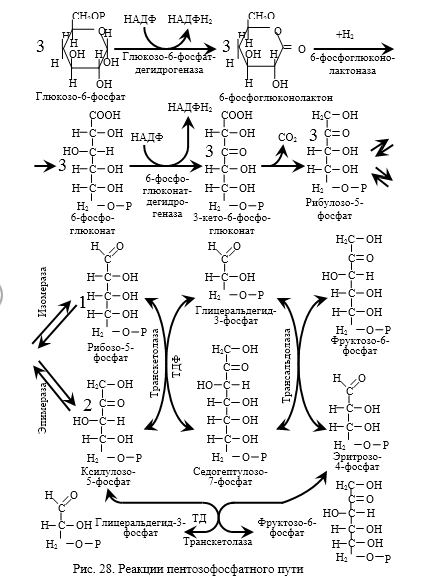 Биологическая роль ПФП ПФП не используется в клетке для наработки АТФ, но выполняет две важнейшие функции. 1. Образование НАДФН2. ПФП более чем на 50 % удовлетворяет потребность тканей в НАДФН2. Молекулы НАДФН2 используются в тканях на следующие цели: а) реакции восстановительного синтеза (синтез жирных кислот, холестерина, стероидных гормонов и т.д.); б) микросомальное окисление ксенобиотиков (реакции гидроксилирования для обезвреживания чужеродных веществ); в) обезвреживание NH3 (реакция восстановительного аминирования). 2. Образование пентоз (рибозо-5-фосфата). ПФП на 100 % удовлетворяет потребности тканей в рибозе, т.е. ПФП является единственным путем, в котором синтезируется рибоза в организме. Рибоза используется для наработки мононуклеотидов, которые в свою очередь идут на синтез многих важнейших веществ:  Путь глюкуроновой кислоты. Основные реакции, биологическая роль. Эссенциальная пентозурия. Путь глюкуроновой кислоты обеспечивает наработку УДФ-глюкуроновой кислоты для синтеза гликозаминогликанов, а также для реакций конъюгации при детоксикации (обезвреживании) и инактивации веществ в печени. С использованием глюкуроновой кислоты в печени происходит конъюгация стероидных гормонов, билирубина и многих лекарств. В результате увеличивается растворимость гидрофобных веществ и облегчается их выведение из организма с мочой. Схема пути глюкуроновой кислоты представлена на рис. 38. 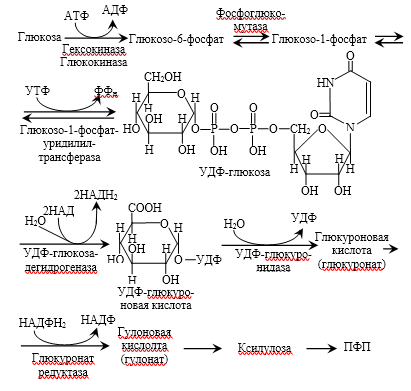 Роль Глюкуронового пути Значение глюкуронового пути: 1.Образование активированного глюкуроната. В гепатоцитах УДФ-глюкуроновая кислота используется на процессы обезвреживания (реакции конъюгации с билирубином, продуктами гниения белков, лекарствами и др.). В фибробластах УДФ-глюкуроновая кислота используется на синтез гетерополи- сахаридов (гиалуроновая кислота, хондроитинсульфат, дерматансульфат, гепарин). 2.Дополнительный источник пентоз. 3.Путь включения пищевого ксилитола в метаболизм. 4.Поставляет гулоновую кислоту на синтез аскорбата. Аскорбат синтезируется из гулоновой кислоты с участием двух специфических ферментов. Один из этих ферментов отсутствует у человека (отсутствует также у высших приматов, морской свинки, индийской летучей мыши), поэтому аскорбат не синтезируется и должен поступать с пищей. Эссенциальная фруктозурия Болезнь отличается доброкачественным течением и отсутствием клинических признаков. Наследуется аутосомно-рецессивно и вызвана отсутствием в печени энзима фруктокиназы. Биохимически устанавливается фруктоземия (40-80 мг% фруктозы в крови) и фруктозурия (с мочой выделяется 10-20% принятой фруктозы). При нагрузке фруктозой гипогликемия не наступает; этим доброкачественная эссенциальная фруктозурия отличается от непереносимости фруктозы. Является доброкачественным, протекающим без клинических признаков, нарушением обмена L-ксилулозы (пенюзы) в результате аутосомно-рецессивно наследуемого дефицита L-ксилулозередуктазы. При отсутствии этого энзима L-ксилулоза скопляется в крови и выделяется с мочой в количестве 2-4 г в сутки. Лечение не является необходимым. |