1. Биология наука о живых системах, закономерностях и механизмах их возникновения, существования и развития. Предмет биологии. Биологические науки, их задачи, объекты изучения. Значение биологии как базисной дисциплины в подготовке врача
 Скачать 4.89 Mb. Скачать 4.89 Mb.
|

14.2.2. Мышечная системаУ представителей типа Хордовые мускулатура подразделяется по характеру развития и иннервации на соматическую и висцеральную. Соматическая мускулатура развивается из миотомов и иннервируется нервами, волокна которых выходят из спинного мозга в составе брюшных корешков спинномозговых нервов. Висцеральная мускулатура развивается из других участков мезодермы и иннервируется нервами вегетативной нервной системы. Вся соматическая мускулатура поперечно-полосата, а висцеральная может быть как поперечно-полосатой, так и гладкой (рис. 14.17). 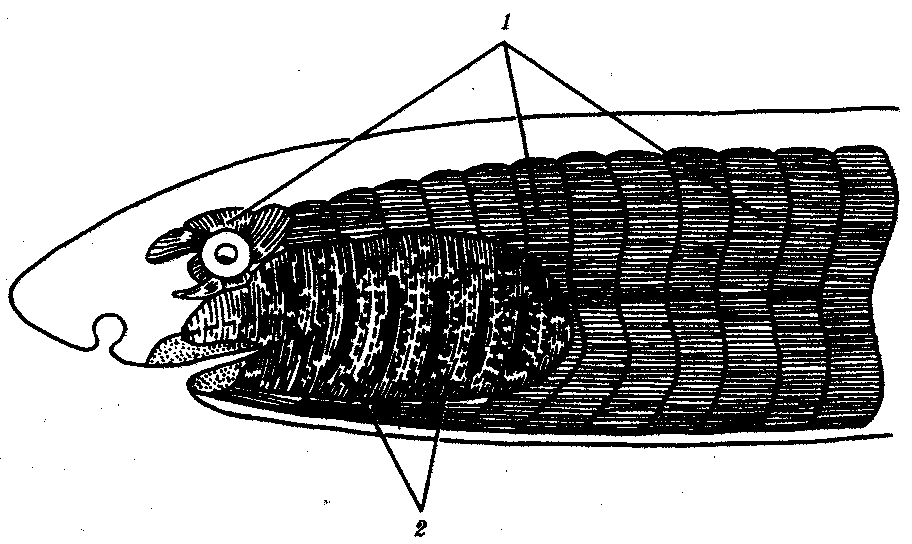 Рис. 14.17. Соматическая и висцеральная мускулатура позвоночных: 1—соматическая мускулатура, развивающаяся из миотомов, 2—висцеральная мускулатура жаберной области 14.2.2.1. Висцеральная мускулатураНаиболее существенные изменения претерпела висцеральная мускулатура, связанная с висцеральными дугами передней части пищеварительной трубки. У низших позвоночных большая часть этой мускулатуры представлена общим сжимателем висцерального аппарата — m. constrictor superficialis, покрывающим всю область жаберных дуг со всех сторон. В области челюстной дуги эта мышца иннервируется тройничным нервом (V), в области подъязычной дуги — лицевым (VII), в области первой жаберной дуги — языкоглоточным (IX), наконец, ее часть, лежащая каудальнее,— блуждающим нервом (X). В связи с этим все производные соответствующих висцеральных дуг и мышц, связанных с ними, иннервируются впоследствии у всех позвоночных перечисленными нервами. В передней части сжимателя обособляется крупная мускулатурная масса, обслуживающая челюстной аппарат. Позади висцерального аппарата дифференцируется трапециевидная мышца m. trapezius, отдельными пучками прикрепляющаяся к последним жаберным щелям и переднему краю спинного отдела плечевого пояса. Часть поверхностного сжимателя в области подъязычной дуги у пресмыкающихся разрастается, охватывает шею снизу и с боков и образует сжиматель шеи m. sphincter colli. У млекопитающих эта мышца делится на два слоя: глубокий и поверхностный. Глубокий сохраняет прежнее название, а поверхностный называется platysma myoides и располагается подкожно. Эти две мышцы разрастаются на всю область головы и дают начало сложной системе лицевой подкожной мускулатуры, которую у приматов и человека называют мимической. Поэтому вся мимическая мускулатура иннервируется так же, как и мышца, из которой она происходит,— лицевым нервом. Мышцы собственно жаберного аппарата с утратой жаберного дыхания редуцируются, но отдельные их элементы сохраняются в виде мышц подъязычного аппарата, глотки и гортани. Трапециевидная мышца полностью теряет связь с жаберным аппаратом и становится исключительно мышцей плечевого пояса. Часть ее у млекопитающих отходит от сосцевидного отростка черепа и приклепляется к ключице и грудине, обособляется — грудино-ключично-сосцевидная мышца т. sternocleidomastoideus. Иннервирующая эту мышцу задняя часть блуждающего нерва становится самостоятельным черепно-мозговым нервом XI пары, добавочным — п. accessorius. Основные этапы филогенеза висцеральной мускулатуры жаберной области рекапитулируют в эмбриогенезе млекопитающих и человека. Знание этих рекапитуляции позволяет объяснить сложность иннервации мышц лица и шеи, объединенных с ними общностью происхождения. 14.2.2.2. Соматическая мускулатураМускулатура головы. У всех позвоночных животных в процессе эмбриогенеза нижние концы миотомов образуют выросты в вентральном направлений, охватывающие полость тела снаружи и срастающиеся по центральной линии на брюшной стороне. Таким образом, закладки соматической мускулатуры становятся сегментарными не только на дорсальной стороне в связи с сегментацией сомитов, но и на вентральной. В миотомах и в их вентральных отростках идет образование продольных мускульных волокон. Миотомы, лежащие на головном конце тела, распадаются на мезенхиму и образуют зачатки отдельных мышц. Из первого миотома головы формируются верхняя внутренняя и нижняя прямые и нижняя косая мышцы глаза, иннервируемые глазодвигательнъш нервом п. oculomotorius (III пара). Из второго миотома — верхняя косая мышца, иннервируемая блоковым нервом п. trochlearis (IV пара); а из третьего — наружная прямая мышца, получающая иннервацию от отводящего нерва п. abducens (VI пара). Задние миотомы головы, образующие мощные вентральные отростки, распространяющиеся вокруг висцеральной мускулатуры глоточной области, образуют подъязычную мускулатуру, которая у рыб слабо дифференцирована, а начиная с земноводных распадается на m. sternohyoideus, т. omohyoideus и т. geniohyoideus. У наземных позвоночных за счет последней мышцы формируется собственная мускулатура языка — m. genioglossus и m. hyoglossus. Вся подъязычная мускулатура иннервируется подъязычным нервом п. hypoglossus, который у амниот становится типичным черепно-мозговым нервом. Мускулатура туловища и конечностей. У бесчерепных, а также у рыб вся мускулатура туловища состоит из ряда мышечных сегментов, или миомеров, правой и левой сторон, которые вместе образуют так называемые боковые мышцы. Каждый миомер развивается из миотома одного сомита и иннервируется первоначально двигательной ветвью одного спинномозгового нерва. Миомеры отделены друг от друга миосептами — соединительнотканными перегородками. Такой же перегородкой, идущей продольно, бокойая мышца разделена на спинную и брюшную мышцы. Уже у рыб направление пучков мышечных волокон в миомерах начинает изменяться на разной глубине мышечного слоя. Эта дифференцировка значительно более выражена у наземных позвоночных и приводит у них к постепенному обособлению различных слоев брюшной и спинной мускулатуры. В результате этого возникают сложные группы мышц, от первоначальной четкой метамерии которых остаются только следы в виде глубоких мышц спины и шеи, связывающих друг с другом соседние позвонки. Значение спинной мускулатуры наземных позвоночных снижается в связи с передвижением большинства из них с помощью конечностей, а брюшные мышцы претерпевают смену функций: первоначально принимающие участие в перемещениях тела, они у пресмыкающихся и млекопитающих служат для изменения объема грудной и брюшной полостей в процессе дыхания. Мускулатура парных плавников рыб закладывается в виде ряда мускульных почек, вырастающих от вентральных концов миотомов. Каждая из этих почек подразделяется на два мышечных зачатка, врастающих в основание закладки плавника с его спинной и брюшной сторон. Первые функционально становятся мышцами, отводящими плавник, вторые — приводящими. У наземных позвоночных из мышечного зачатка, гомологичного отводящей мышце плавника, развивается группа разгибателей пятипалой конечности, а из зачатка ее антагонистов — группа сгибателей. В пределах каждой группы идет дифференцировка на поверхностные и глубокие мышечные пучки, становящиеся самостоятельными мышцами. В целом мышцы наземных позвоночных, гомологичные мышцам плавников рыб, образуют первичную мускулатуру конечностей. Она иннервируется нервами плечевого и пояснично-крестцового сплетений, образованных в процессе перемещения поясов конечностей в ходе эмбриогенеза (см. разд. 14.2.1). При дальнейшей дифференцировке миотомов грудной клетки развивается группа мышц, приводящая в движение сам плечевой пояс, или вторичная мускулатура. К ней относят широчайшую мышцу спины, большую и малую грудные, а также зубчатую мышцы. Они иннервируются непосредственно спинномозговыми нервами сегментов спинного мозга, расположенных каудальнее тех его участков, которые осуществляют иннервацию первичной мускулатуры. Задняя пара конечностей не имеет вторичной мускулатуры в связи с тем, что гетеротопия тазового пояса по отношению к позвоночнику в процессе эволюции менее выражена. Изменение среды обитания и характера движений позвоночных привело к усилению и обособлению большого числа мышц, обслуживающих конечности, и к относительной редукции собственно мышц туловища. Такие мышцы, как грудные, широчайшая мышца спины и трапециевидная, в значительной мере покрывают туловищную мускулатуру и даже частично вытесняют ее функционально. 105.Прогрессивный характер эволюции живой материи и возникновение человека. Место человека в системе животного мира. Качественные особенности человека. 106.Биологические предпосылки прогрессивного развития гоминид. Антропогенез. Характеристика основных этапов. Долгое время многие палеонтологи и антропологи считали наиболее древними представителями семейства гоминид южно-африканских двуногих приматов Australopithecusafricanus. Возраст большей части южно-африканских находок австралопитеков определяется в интервале 3—1 млн. лет. Строение скелетов этих организмов свидетельствует об их прямохождении. Масса мозга — 450—550 г при общей массе тела от 25 до 65 кг. Зубная система близка к человеческой: клыки малых 146 размеров, зубы расположены в виде широкой дуги, как у человека. Это свидетельствует о всеядности. В местах обнаружения остатков австралопитеков имеется множество костей, расколотых тяжелыми предметами. Большое число черепов животных расколоты с левой стороны, и это свидетельствует о том, что австралопитеки были в основном правшами. Некоторые австралопитеки, видимо, начинали осваивать огонь. В 1960—1970-х гг. в результате раскопок в Эфиопии и позже в Танзании и Кении были обнаружены большие группы гоминид, древность которых определяется в 4—2,8 млн. лет. По этим материалам в 1978 г. был описан новый вид австралопитеков A. afarensis, более примитивный, чем A. africanus, но тоже двуногий и с чертами «промежуточности» по отношению к современным человекообразным обезьянам и человеку (рис. 15.4). Объем мозговой полости черепа оказался в пределах 380—450 см3, т.е. практически не отличался от объема мозга современного шимпанзе. Действительно, австалопитеки имеют большее сходство с человеком не по объему и строению мозга, а по строению зубной системы и способу передвижения. Изучение австралопитековых показало, что именно двуногость, а не большой объем мозга, явилась ключевой адаптацией ранних гоминид. В 1964 г. по находкам, сделанным в Танзании, был выделен вид Homohabilis, или Человек умелый, имеющий абсолютный возраст 2—1,7 млн. лет. Отличительными чертами его являются двуногость, в целом прогрессивное строение кисти, зубной системы, объем мозговой коробки от 540 до 700 см3, что примерно в полтора раза превышает объем мозга австралопитеков. На внутренней поверхности черепа обнаруживаются признаки прогрессивных нейроморфологических изменений, определяющиеся по отпечаткам головного мозга: выраженная асимметрия полушарий и развитие двух речевых центров как условие для возникновения членораздельной речи. Большой палец стопы не отведен в сторону. Это свидетельствует о том, что морфологические перестройки, связанные с прямохождением, у него полностью завершились. Вместе с остатками Н. habilis найдены орудия труда со следами целенаправленной обработки, свидетельствующие о ранних формах трудовой деятельности. Перечисленные признаки, ведущим из которых является прогрессивное развитие мозга, характеризуют организм уже с иной морфофункциональной организацией по сравнению с австралопитеками. Эти признаки считаются специфичными для рода Homo. Сопоставление морфологии африканского и афарского австралопитеков с Н. habilis и современным человеком позволяет предположить следующую схему филогенетических взаимоотношений ранних гоминид (рис. 15.5). Африканский австралопитек, считавшийся раньше общим предком как рода Homo, так и других австралопитековых, является в этой схеме представителем боковой ветки эволюции, приведшей к узкой специализации и образованию мощных форм типа A. robustus, которые вымерли около 1 млн. лет назад. Общим предком всех австралопитековых и рода Homo является в соответствии с этой схемой A. afarensis. Таким образом, на протяжении 1—1,5 млн. лет представители двух близких родов и, возможно, нескольких видов семейства гоминид сосуществовали, причем не только во времени, но и на перекрывающихся территориях. В основе дивергенции различных линий ранних гоминид и австралопитековых могли лежать разного рода механизмы изоляции, в первую очередь генетические: мутации в виде хромосомных перестроек. Это означает, что эволюция австралопитековых шла постепенно, приводя благодаря дивергенции к морфологическому и экологическому разнообразию. Ведущими факторами эволюции на прегоминидной стадии антропогенеза являлись, несомненно, факторы биологической эволюции, главным из которых является естественный отбор. Об этом свидетельствует, в частности, большое видовое разнообразие австралопитековых, обитавших в различных условиях практически на всей территории Южной, Центральной и Северо-Восточной Африки. В это же время в происхождении рода Homo имело место скачкообразное изменение наследственного материала. В разных органах и системах прегоминид обнаруживалась асинхронность филогенеза. Есть предположение о том, что эволюция коры больших полушарий мозга состоит из двух компонентов, разобщенных по времени: соматического, обеспечивающего сенсорно-моторные функции, и несоматического, связанного с высшими психическими функциями. Если локомоторный комплекс подвергался длительным постоянным изменениям, то головной мозг эволюционировал скачкообразно. Элементы скачкообразности в эволюции некоторых структур ранних гоминид могли быть обусловлены «транспозиционными взрывами» (перестройками генома за счет подвижных генетических элементов, переносящих комплексы генов на новое место). Это могло сопровождаться постепенным развитием других морфофизиологических свойств в результате накопления малых мутаций под контролем естественного отбора. Но именно в период становления Н. habilisвозникла, вероятно, часть хромосомных перестроек в геноме человека, о которых говорилось выше. Следующей ступенью гоминизации после появления Н. habilisсчитается возникновение архантропов, представителем которых является сборный вид Н. erectus, или Человек прямоходящий. Материальная культура и ярко выраженная социальность позволили ему быстро и эффективно расселиться по всей территории Африки и Евразии и освоить обширный ареал, разнообразный в климатическом отношении. Действительно, орудия труда Н. erectus более прогрессивны, чем у Н. habilis, а масса мозга (от 800 до 1000 г) явно превышает минимальную массу (750 г), при которой возможно существование речи. Наличие при этом речевых центров, возникших впервые у Н. habilis, предполагает и развитие второй сигнальной системы. Выделяют три группы Н. erectus: обитавшие в Европе, Азии и Африке. Долгое время древнейшими архантропами считались азиатские представители из Индонезии и Восточного Китая — питекантроп и синантроп. Однако находки последних лет на территории Израиля (1982) и Кении (1984), датирующиеся соответственно 2,0 и 1,6 млн. лет, сопровождающиеся элементами материальной культуры и признаками использования огня, показали, что арогенная эволюция гоминид происходила на африканском континенте и на Ближнем Востоке. Это позволило связать происхождение Н. erectus с восточноафриканскими формами Н. habilis. Наличие большого количества находок архантропов древностью 1,5—0,1 млн. лет в отдаленных от Африки регионах — в Юго-Восточной и Восточной Азии, в Центральной Европе и даже на Британских о-вах — свидетельствуют об активных адаптациях их к разнообразным условиям существования (рис. 15.6). В связи с тем что небольшое различие ископаемых остатков Н. erectus не соответствует значительному разнообразию природно-климатических условий указанных территорий, можно заключить, что в этих адаптациях значительную роль играли наряду с факторами биологической эволюции также и социальные факторы: совместное изготовление укрытий, орудий труда и использование огня. Роль Н. erectus как этапа антропогенеза никогда не подвергалась сомнению. Что же касается палеоантропа, или неандертальского человека, то его роль в происхождении человека в настоящее время оспаривается. Это связано в первую очередь с обнаружением большого количества ископаемых остатков человека с промежуточными чертами между Н. erectus и человеком современного физического типа. Кроме того, палеонтологические находки последних лет дают повод судить о недооценке интеллектуальных возможностей неандертальцев. На всех стоянках обнаружены следы костров и обгоревшие кости животных, что свидетельствует об использовании огня для приготовления пищи. Орудия труда их гораздо совершеннее, чем у предковых форм. Масса мозга неандертальцев около 1500 г, причем сильное развитие получили отделы, связанные с логическим мышлением. Костные остатки неандертальца из Сен-Сезер (Франция) были найдены вместе с орудиями труда, свойственными верхнепалеолитическому человеку, что свидетельствует об отсутствии резкой интеллектуальной грани между неандертальцем и современным человеком. Имеются данные о ритуальных захоронениях неандертальцев на территории Ближнего Востока. Эти и ряд других находок позволили в конце 60-х годов выделить палеоантропов в отдельный подвид Homosapiensneanderthalensis в отличие от неоантропа H.s. sapiens, который, таким образом, тоже получил ранг подвида. Наиболее древние ископаемые остатки его возрастом 100 тыс. лет обнаружены также на территории Северо-Восточной Африки. Многочисленные находки палеоантропов и неоантропов на территории Европы, датирующиеся 37—25 тыс. лет, свидетельствуют о существовании обоих подвидов в течение нескольких тысячелетий. В тот же период неоантропы обитали уже не только в Европе и Африке, но и в отдаленных районах Азии (о. Тайвань, о. Окинава) и даже в Америке. Эти данные указывают на необычайно быстрый процесс расселения современного человека, что может быть доказательством «взрывного», скачкообразного характера антропогенеза в этот период как в биологическом, так и в социальном смысле. H.s. neanderthalensis в виде ископаемых остатков не обнаруживается позже рубежа в 25 тыс. лет. Быстрое исчезновение палеоантропов может быть объяснено вытеснением их людьми с более совершенной техникой изготовления орудий труда и метисацией с ними. С возникновением человека современного физического типа роль биологических факторов в его эволюции свелась к минимуму, уступив место социальной эволюции. Об этом отчетливо свидетельствует отсутствие существенных различий между ископаемым человеком, жившим 30—25 тыс. лет назад, и нашим современником. 107.Соотношение биологических и социальных факторов в становлении человека на разных этапах антропогенеза. Биологическое и социальное наследование человека современного физического типа. Возрастающая роль социального наследования. В человеке 2 компонента: биологический (соответствует биологическому прошлому) и социальный (соответствует настоящему). Для того чтобы человек стал социальным существом нужны предпосылки, в первую очередь, морфологические. Предшественники человека – древесные формы. Млекопитающие, живущие на деревьях, точно определяют расстояние до предмета, удерживаются на ветках, фокусируют взгляд на предмете, кроме того, обладают способностью хватать предметы, перелетая с ветки на ветку. Приматы имеют хорошее зрение. Глаза хорошо развиты, направлены вперед, происходит фокусировка глаз на одном предмете и определение расстояния. Наличие желтого пятна еще лучше делает зрение, цветное зрение, улучшает объемное видение. Зрение – значительный компонент ВНД. От сочетания глаз/рука зависит развитие мозга. Концентрирование на одном предмете осуществляется именно сочетанием глаз/рука. Эта способность легла в основу экспериментирования, изготовления орудий и решений задач. Передвижение на 2 ногах освободило руки, которые стали использовать орудия и их изготавливать. Труд – решающий шаг в выделении человека из животного мира. Маркс и Энгельс писали: «Труд – первое основное условие всей человеческой жизни и притом в такой степени, что мы можем сказать в известной степени, что труд создал человека». Свободная рука была вначале подходящая для простых действий, она долго развивалась. Труд оказывал прямое влияние на строение и функции руки, изменения передавались по наследству. Под влиянием труда кисть формируется и изменяется в определенном направлении. В строении и функции руки отражена трудовая деятельность в целом. Рука – часть тела, которая, изменяясь, оказывала влияние на организм, причем организм в свою очередь влиял и на развитие руки. Трудовая деятельность повлекла улучшение не только физической, но и умственной организации, происходила цефализация. Развитие головного мозга имело огромную приспособительную ценность. Масса головного мозга еще ни о чем не говорит. Необходимо улучшить строение мозга, увеличиваются лобная, теменная, височная доли. После развития мозга никаких значительных морфологических изменений не было, за исключением удлинения ног. Морфологически человек не изменился. «Древние люди – готовые люди» - Энгельс. Морфологический прогресс человека завершился на самых ранних этапах развития. Но биологический процесс не завершился. С появлением древнейших людей появляются зачатки трудовой деятельности, общество. Труд требовал передачи опыта и объединений усилий. В процессе общественного труда возник звуковой язык, а затем – членораздельная речь. Речь могла возникнуть только тогда, когда мозг был достаточно хорошо развит. 750 г – критическая масса мозга, начиная с которой человек обладает предпосылками для овладения речью и языком. Необходимы специализированные нервные окончания, механизмы, управляющие дыханием, колебанием голосовых связок с определенной частотой. Человек отличается от млекопитающих т.к. они приспособлены к модулированию звуков. У приматов имеется надгортанник и мягкое небо, поэтому они не задыхаются. Младенцы способны есть и дышать одновременно. Язык не имеет достаточного места для движения и артикуляции звуков речи – это у младенцев. У взрослых гортань не входит в носовую полость, пища с питьем проходят мимо гортани, вход в которую плотно закрывается мышцами – защитные движения. Человек может задохнуться, языку места достаточно. 2000 людей в США ежегодно умирают от удушья. В эмбриогенезе гортань сначала располагается высоко. Затем опускается. К 3 месяцам после рождения ребенок может дышать через рот. Труд и речь оказывали обратное мощное влияние на прогрессивное развитие органов чувств и головного мозга. Под влиянием трудовой деятельности естественный набор изменил направление эволюции. Выживали те, кто лучше охотился, изготавливал орудия, заботился о стариках. Люди все меньше завесили от внешней среды. Если бы предки человека не занялись трудом – они бы вымерли. Прямохождение имеет отрицательное значение: медленное передвижение, не выгодно энергетически. Роды у женщин мучительны, а у четвероногих – нет т.к. крестец у человека лишается подвижности. Длительное стояние, ношения тяжестей приводит к плоскостопию, расширению вен. Органы давят друг на друга и на таз, так возникают грыжи, аппендицит, опущение и выпадение матки. Человек занял главное место в животном мире. Его преимущества – свободная рука и хорошо развитый головной мозг. Возникновение и развитие социальной информации. Биологические закономерности постепенно заменялись социальными. Произошло превращение зоопсихологической информации в социальную в процессе рудовой деятельности. Социальная информация возникла из ненаследственной части зоопсихологической информации и превратилась в наследуему3ю. В.М.Бехтерев в 1927 году «Есть особый способ передачи приобретенных способностей потомству. Этот способ мы называем социальной наследственностью». Дубинин развивал эту идею. « Наряду с генетической программой люди создали социальную». Важная часть эволюции человека – содержание социальной программы генетически не закрепляется. Результаты трудовой и общественной деятельности передавались из поколения в поколение путем воспитания. Труд, воля, интеллект – это отражение человеком исторически сложившихся отношений. В каждом поколении каждый человек сам на основе генетической информации проходит заново формирование. Сущность человека – биосоциальная. Биологическая сущность – предпосылка социальной. Нарушение структуры ДНК приводит к тому, что биологические особенности не отвечают социальным требованиям. И биологическое и социальное должно быть в норме, тогда возникает гармоничная личность. Если ребенок лишен общества, у него нет речи, сознания. «Дикие дети». Социальные свойства человека развиваются в общении с другими людьми. 1980 год – слепоглухонемые дети от рождения. Ранее они были вне общества, но Мещеряков и Серебрянский добились своими методами появления сознания и речи. Прекращение видообразования человека разумного. Эволюция не происходит, следовательно, естественного отбора нет. Но завершенность видообразования еще не означает завершения эволюционного процесса, как это кажется на первый взгляд. Просто движущая форма отбора сменилась стабилизирующей. Поддержание физиологического типа современного человека с возникновением трудовой деятельности и социальной информации завершается действие группового отбора как главного двигателя эволюции. Но биологические факторы эволюции были устранены не сразу. Появился общебиологический процесс, социальной стало главенствующим. Человек – существо социальное. Человек приспосабливал природу для своих нужд. Он почти перестал зависеть от природы. Серебровский « Если ход эволюции есть постепенное освобождение от природы, то он сам должен воздействовать на природу» 108.Внутривидовая дифференциация человечества. Расы и расогенез. Видовое единство человечества. Современная классификация и распространение человеческих рас. Популяционная концепция рас. С момента возникновения Н. sapiens социальное в человеке стало его сущностью и биологическая эволюция видоизменялась, проявляясь в возникновении широкого генетического полиморфизма (см. § 12.3). Генетическое разнообразие на уровне генов и в меньшей степени хромосом обеспечивает разнообразие генотипов особей. Разнообразные генотипы по-разному проявляются в меняющихся условиях среды, давая огромное фенотипическое многообразие людей. В основе морфофизиологического полиморфизма человечества лежат полиморфизм наследственного материала на уровне генома и модификационная изменчивость. Эти факторы обеспечивают не только индивидуальное морфофизиологическое многообразие, но и внутривидовую групповую дифференциацию человечества на расы и адаптивные экологические типы. |