Шпора. Экзамен по биологии. 1. Биология. Жизнь, происхождение. Уровни организации жизни. Термин биология
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Взаимодействие аллельных генов. Постоянно возникающие разнообразные изменения структуры генов обусловливают явление множественного аллелизма, поэтому взаимодействующие при оплодотворении гаметы часто несут в своих геномах разные аллели одного и того же гена. В связи с этим генотип нового организма является гетерозиготным по многим локусам, т.е. его аллельные гены, расположенные в соответствующих участках гомологичных хромосом и пришедшие от обоих родителей, представлены разными аллелями— А и А', В и В' С и С' Если аллельные гены представлены одинаковыми аллелями, т.е. находятся в гомозиготном состоянии, то развивается соответствующий данному аллелю вариант признака. В случае гетерозиготности развитие данного признака будет зависеть от взаимодействия аллельных генов. Доминирование Называется тип взаимодействия аллейных генов, при котором один ( доминантный) аллейный ген у гибридов первого поколения полностью подавляет проявление другого (рецессивного) аллейного гена. Примером доминирования одного из аллелей в гетерозиготном генотипе может служить определение групповой принадлежности крови у человека по системе АВ0. Неполное доминирование Наблюдается, когда фенотип гетерозигот BB' отличается от фенотипа гомозигот по обоим аллелям (BB или B'B') Промежуточным проявлением признака. Это объясняется тем, что аллель, способный сформировать нормальный признак, находясь в двойной дозе у гомозиготы BB, проявляется сильнее, чем в единственной дозе у гетерозиготы BB'. Указанные генотипы отличаются экспрессивностью, т.е. степенью выраженности признака. Демонстрацией такого типа взаимодействия генов могут быть многочисленные наследственные заболевания у человека, проявляющиеся клинически у гетерозигот по мутантным аллелям, а у гомозигот заканчивающиеся смертью. Иногда гетерозиготы имеют почти нормальный фенотип, а гомозиготы характеризуются пониженной жизнеспособностью. Так, гомозиготы по аллелю серповидноклеточности эритроцитов в связи с развитием у них тяжелой формы анемии и других фенотипических проявлений обычно не переживают детский возраст. Напротив, гетерозиготы—это как правило нормальные люди. Кодоминирование Представляет собой такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате этого формируется некий промежуточный вариант признака, новый по сравнению с вариантами, определяемыми каждым аллелем самостоятельно. Примером может служить формирование IV, или АВ-группы, крови у человека, гетерозиготного по аллелям IA иIB, которые по отдельности детерминируют образование II и III групп крови. Межаллельная комплементация Относится к достаточно редко встречаемым способам взаимодействия аллельных генов. В этом случае возможно формирование нормального признака D у организма, гетерозиготного по двум мутантным аллелям гена D(D'D"). Допустим, что ген D отвечает за синтез какого-то белка, который имеет четвертичную структуру, состоящую из нескольких одинаковых пептидных цепей. Мутантный аллель D'определяет синтез измененного пептида D', a мутантный аллель D" приводит к синтезу другой, но тоже измененной структуры пептида D". Можно представить ситуацию, когда взаимодействие таких измененных пептидов (D' и D") при формировании четвертичной структуры, как бы взаимно компенсируя эти изменения, обеспечивает образование белка с нормальными свойствами. В то же время отдельно взаимодействующие пептиды D' или D" формируют аномальные белки. Таким образом, с определенной вероятностью у гетерозигот D'D" в результате межаллельной комплементации может образовываться нормальный признак в виде белка с нормальными свойствами. Аллельное исключение Такой вид взаимодействия аллельных генов в генотипе организма, который можно понять на примере рассмотренного выше механизма инактивации одной из Х-хромосом у особей гомогаметного пола, приводящего всоответствие дозы Х-генов у всех представителей вида. Инактивация одного из аллелей в составе Х-хромосомы способствует тому, что в разных клетках организма, мозаичных по функционирующей хромосоме, фенотипически проявляются разные. Аллельное исключение наблюдается также в В-лимфоцитах, синтезирующих специфичные антитела к определенным антигенам. Моноспецифичность таких иммуноглобулинов требует выбора, который должна осуществить каждая клетка между экспрессией отцовского или материнского аллеля. 25. Закон независимого наследования признаков и его цитологическое обоснование. Примеры независимого наследования признаков у человека. "При дигибридном скрещивании двух гетерозиготных особей каждая пара признаков наследуется независимо от другой и дает с ним различные сочетания." Этот закон действует в том случае, если гены, контролирующие разные признаки, лежат в разных парах хромосом. Гибриды могут дать 4 типа гамет. Открытие независимого характера наследования разных признаков у гороха дало возможность Менделю высказать предположение о дискретности наследственного материала, в котором за каждый признак отвечает своя пара наследственных задатков, сохраняющих в ряду поколений свою структуру и не смешивающихся друг с другом. Современные представления о надмолекулярной организации наследственного материала в хромосомах и закономерностях их передачи в ряду поколений клеток и организмов объясняют независимый характер наследования признаков расположением соответствующих генов в негомологичных хромосомах. Примеры независимого наследования признаков у человека. Шизофрения, Бронхиальная астма, Расщелина губы и нёба, Сердечно-сосудистые заболевания, Артериальная гипертензия, Дефекты невральной трубки, Изолированный врожденный порок сердца. Чем сильнее выражен признак или тяжелее протекает заболевание у больного родственника, тем выше риск его развития у здоровых родственников. 26. Виды взаимодействия неаллельных генов. Примеры у человека. Взаимодействие неаллельных генов Неаллельные гены – гены, расположенные в различных участках хромосом и кодирующие разные белки 1) Комплементарность – процесс взаимодействия двух и более взаимодополняющих неаллельных генов, при котором появление признака у человека обусловливается обязательным присутствием этих генов в определенном состоянии ( обычно доминантном) Пример развитие нормального слуха определяют два гена: А – отвечает за нормальное развитие улитки, и В – отвечает за нормальное развитие слухового нерва. Поэтому если один из генов в гомозиготном рецессивном состоянии, то человек будет глухим. 2)Эписта́з — взаимодействие генов, при котором активность одного гена находится под влиянием другого гена (генов), неаллельного ему. Ген, подавляющий фенотипические проявления другого, называется эпистатичным; ген, чья активность изменена или подавлена, называется гипостатичным. У человека примером доминантного эпистаза являются ферментопатии(энзимопатии) – заболевания, в основе которых лежит недостаточная выработка того или иного фермента. Пример рецессивного эпистаза – так называемый «бомбейский феномен»: в семье у родителей, где мать имела группу крови О, а отец – группу крови А, родились две дочери, из которых одна имела группу крови АВ. Ученые предположили, что у матери в генотипе был ген IB, однако его действие было подавлено двумя рецессивными эпистатическими генами dd 3) Полимери́я — взаимодействие неаллельных множественных генов, однонаправленно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопи тельной) полимерии степень проявления признака зависит от суммарного действия нескольких генов. Чем больше доминантных алле лей генов, тем сильнее выражен тот или иной признак. Расщепле ние в F2 по фенотипу при дигибридном скрещивании происходит в соотношении 1:4:6:4:1, а в целом соответствует третьей, четвертой (при дигибридном скрещивании), седьмой (при тригибридном скрещивании) и т.п. строчкам в треугольнике Паскаля. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление в F2 по фенотипу при дигибридном скрещивании — 15:1. Пример полимерии — наследование цвета кожи у людей, который зависит (в первом приближении) от четырёх генов с кумулятивным эффектом. 27. Сцепленное наследование генов. Группы сцепления. Карты хромосом и методы их со ставления. Сцепленное наследование генов. Группы сцепления. Карты хромосом и методы их составления. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом. Генетической картой хромосомы называется схема относительного расположения генов, входящих в состав одной хромосомы и принадлежащих к одной группе сцепления. Для составления хромосомной карты необходимо определить число групп сцепления, затем принадлежность гена к той или иной группе сцепления и, наконец, расположение гена в хромосоме по отношению к другим генам. На первом этапе картирования определяют принадлежность гена к той или иной группе сцепления. Как известно, у D. melanogaster в диплоидном наборе четыре пары хромосом: первая пара — половые хромосомы (XX — у самок, XY — у самцов), вторая, третья и четвертая — аутосомы. Число генов в Y-хромосоме самцов очень мало. Для локализации вновь возникшей мутации необходимо располагать набором маркерных генов для каждой хромосомы. Картирование мутации основывается на анализе ее сцепления с этими маркерами. Например, если интересующая нас мутация наследуется независимо от маркеров второй хромосомы, делается вывод о ее принадлежности к другой группе сцепления. Скрещивания проводятся до тех пор, пока не удастся выявить сцепленное наследование анализируемой мутации с маркерными мутациями какой-либо хромосомы. Второй этап картирования подразумевает определение положения гена на хромосоме. Для этого подсчитывают расстояние между этим геном и уже известными, маркерными генами. Для подсчета генетических расстояний проводят специальные скрещивания, в потомстве которых учитывают частоты кроссоверных и некроссоверных особей. Предполагается, что расстояние между двумя генами пропорционально частоте кроссинговера между ними. Следует иметь в виду, что, чем дальше расположены друг от друга гены, тем чаще между ними происходят множественные перекресты и тем больше искажается истинное расстояние между этими генами. Частая рекомбинация между расположенными далеко друг от друга генами может привести к увеличению числа кроссоверных организмов в потомстве анализирующего скрещивания до 50%, имитируя независимое наследование изучаемых признаков. Поэтому при составлении карт расстояния между далеко расположенными генами следует использовать не непосредственный подсчет числа кроссоверных особей в анализирующих скрещиваниях, а сложение расстояний между многими близко расположенными друг от друга генами, находящимися внутри изучаемого протяженного участка. В этом случае сцепление между далеко расположенными генами можно установить по их сцепленному наследованию с промежуточно-расположенными генами, которые в свою очередь сцеплены между собой. В результате такого метода определения расстояний между генами длины карт хромосом могут превышать 50 морганид. Так, у дрозофилы генетическое расстояние между генами, лежащими в разных концах хромосомы 2, составляет 107 морганид. Метод цитологических карт основан на использовании хромосомных перестроек. При облучении и действии других мутагенов в хромосомах часто наблюдаются потери (делеции) или вставки (дупликации) небольших фрагментов, сравнимых по величине с одним или несколькими локусами. Например, можно использовать гетерозиготы по хромосомам, одна из которых будет нести группу следующих друг за другом доминантных аллелей, а гомологичная ей — группу рецессивных аллелей тех же генов ABCDE/abcde. Если в хромосоме с доминантными генами произошла утрата отдельных генов, например DE, то у гетерозиготы ABC/abcde будут проявляться рецессивные признаки de. На этом принципе основан метод перекрывающихся делеции, используемый при построении цитологических карт. 28.Хромосомная теория наследственности. Сцепленное наследование признаков. Приме ры сцепленного наследования признаков у человека. Хромосомная теория наследственности — теория, согласно которой передача наследственной информации в ряду поколений связана с передачей хромосом, в которых в определённой и линейной последовательности расположены гены. — это совместное наследование генов, расположенных в одной и той же хромосоме. Сцепленное наследование признаков - — это совместное наследование генов, расположенных в одной и той же хромосоме. Примеры при сцеплении с полом дальтонизм, гемофилия, гипертрихоз, раннее облысение, миопатия Дюшенна (Х хромосома) , судорожные расстройства, фенилкетонурия, сахарный диабет (У хр)
29. Группы сцепления. Карты хромосом и методы их составления. Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом. Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел. Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет. Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей. Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей. Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам. У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза). 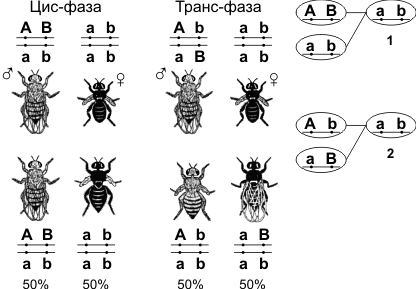 Механизм цис- и транс-фазы 1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы). Картирование хромосом Для составления карт хромосом рассчитывают взаимное расстояние между отдельными парами генов и затем определяют расположение этих генов относительно друг друга. Так, например, если три гена расположены в следующем порядке: А В С, то расстояние между генами А и С (процент рекомбинаций) будет равно сумме расстояний (процентов рекомбинаций) между парами генов АВ и ВС. Если гены расположены в порядке: А С В, то расстояние между генами А и С будет равно разности расстояний между парами генов АВ и СВ. Задача 6-23 Гены А, В и С находятся в одной группе сцепления. Между генами А и В кроссинговер происходит с частотой 7,4%, а между генамиВ и С – с частотой 2,9%. Определить взаиморасположение генов А, В и С, если расстояние между генами А и С равняется 10,3% единиц кроссинговера. Как изменится взаиморасположение этих генов, если частота кроссинговера между генами А и С будет составлять 4,5%? |