пз 10 ост- физиология. 1. Общий план структурнофункциональной организации анализаторов. Анализатор
 Скачать 1.06 Mb. Скачать 1.06 Mb.
|

|
1. Общий план структурно-функциональной организации анализаторов. Анализатор – совокупность центральных и периферических образований, воспринимающих и анализирующих изменения внешней и внутренней сред организма. Сенсорные системы – это специализированные части нервной системы, включающие периферические рецепторы (сенсорные органы, или органы чувств), отходящие от них нервные волокна (проводящие пути) и клетки центральной нервной системы, сгруппированные вместе (сенсорные центры). По И. П. Павлову анализатор имеет три отдела: периферический, проводниковый и центральный, или корковый. 1-Периферический отдел анализатора представлен рецепторами. Его назначение - восприятие и первичный анализ изменений внешней и внутренней сред организма. В рецепторах происходит трансформация энергии раздражителя в нервный импульс, а также усиление сигнала за счет внутренней энергии метаболических процессов. Для рецепторов характерна специфичность (модальность), т.е. способность воспринимать определенный вид раздражителя, к которому они приспособились в процессе эволюции (адекватные раздражители), на чем основан первичный анализ (рецепторы зрительного анализатора - восприятие света, слуховые рецепторы – звук). Та часть рцепторной поверхности, от которой сигнал получает одно афферентное волокно, называется его рецептивным полем. 2-Проводниковый отдел анализатора включает афферентные (периферические) и промежуточные нейроны стволовых и подкорковых структур центральной нервной системы (ЦНС), которые составляют как бы цепь нейронов, находящихся в разных слоях на каждом уровне ЦНС. Проводниковый отдел обеспечивает проведение возбуждения от рецепторов в кору большого мозга и частичную переработку информации. Проведение возбуждения по проводниковому отделу осуществляется двумя афферентными путями: 1) специфическим проекционным путем (прямые афферентные пути) от рецептора по строго обозначенным специфическим путям с переключением на различных уровнях ЦНС 2) неспецифическим путем, с участием ретикулярной формации. На уровне ствола мозга от специфического пути отходят коллатерали к клеткам ретикулярной формации, к которым могут конвергировать различные афферентные возбуждения, обеспечивая взаимодействие 15 анализаторов. Обеспечивает вегетативный, двигательный и эмоциональный компоненты сенсорных реакций. 3-Центральный, или корковый, отдел анализатора, по И.П. Павлову, состоит из двух частей: центральной части, т.е. ≪ядра≫, представленной специфическими нейронами, перерабатывающими афферентную импульсацию от рецепторов, и периферической части, т.е. ≪рассеянных элементов≫ – нейронов, рассредоточенных по коре большого мозга. Корковые концы анализаторов называют также ≪сенсорными зонами≫, которые не являются строго ограниченными участками, они перекрывают друг друга. Рецепторы характеризуются большим разнообразием. В классификации рецепторов центральное место занимает их деление в зависимости от вида воспринимаемого раздражителя. Существует пять типов таких рецепторов. 1. Механорецепторы возбуждаются при их механической деформации. 2. Хеморецепторы воспринимают химические изменения внешней и внутренней среды организма 3. Терморецепторы воспринимают изменения температуры. 4. Фоторецепторы в сетчатке глаза воспринимают световую (электромагнитную) энергию. 5. Ноцицепторы, возбуждение которых сопровождается болевыми ощущениями. 2 Кодирование сенсорной информации Кодирование информации в нервной системе – это преобразование специфической энергии стимула (света, звука, давления и др.) в универсальные коды нейронной активности, на основе которых мозг осуществляет процесс переработки информации. В сенсорной системе сигналы кодируются двоичным кодом, т.е. наличием или отсутствием электрического импульса в тот или иной момент времени Информация о раздражении и его параметрах передается в виде отдельных импульсов, а также групп, или «пачек» импульсов Амплитуда, длительность и форма каждого импульса одинаковы, но количество импульсов в пачке, частота их следования, длительность пачек и интервалов между ними, а также временной «Рисунок» (pattern) пачки различны и зависят от характеристик стимула Например В слуховой сенсорной системе механическое колебание перепонки и других звукопроводящих элементов на первом этапе преобразуется в рецепторный потенциал, последний обеспечивает выделение медиатора в синаптическую щель и возникновение генераторного потенциала, в результате действия которого в афферентном волокне возникает нервный импульс. Потенциал действия достигает следующего нейрона, в синапсе которого электрический сигнал снова превращается в химический, т. е. многократно меняется код. Следует отметить, что на всех уровнях сенсорных систем не происходит восстановления стимула в его первоначальной форме. Этим физиологическое кодирование отличается от большинства технических систем связи, где сообщение, как правило, восстанавливается в первоначальном виде. Пространственное кодирование Пространственное кодирование - информация кодируется количеством и определенной топографией нервных волокон, приносящих возбуждение в ЦНС. 3. Особенности соматовисцеральной сенсорной системы.
4. Тактильный анализатор. Тактильный анализатор является частью кожного анализатора. Они служат для анализа всех механических влияний, действующих на тело человека. Рецепторы, предназначенные для этого, содержатся в коже, в частности, в эпидермисе, дерме и частично в подкожной клетчатке. Концентрация тактильных рецепторов на различных участках тела неодинакова, поэтому чувствительность одних участков выше, например, кожи кончиков пальцев рук, других — ниже. Рецепторы обеспечивает ощущения прикосновения, давления, вибрации и щекотки. Выделяют 3 основных вида рецепторов: 1. Рецепторы давления, которые воспринимают силу механического воздействия (рецепторы силы). 2. Рецепторы прикосновения, или датчики скорости - это тельца Мейсснера. 3. Рецепторы вибрации - это датчики ускорения или датчики синусоидального изменения силы. Они реагируют лишь на вторые производные изменения силы - ускорение. Морфологически они представлены тельцами Паччини. Расположены в глубоких слоях дермы. Периферический отдел представлен различными рецепторными образованиями, раздражение которых приводит к формированию специфических ощущений. На поверхности кожи, лишенной волос, а также на слизистых оболочках на прикосновение реагируют специальные рецепторные клетки (тельца Мейснера), расположенные в сосочковом слое кожи. На коже, покрытой волосами, на прикосновение реагируют рецепторы волосяного фолликула, обладающие умеренной адаптацией. На давление реагируют рецепторные образования (диски Меркедя), расположенные небольшими группами в глубоких слоях кожи и слизистых оболочек. Это медленно адаптирующиеся рецепторы. Адекватным стимулом для них служит прогибание эпидермиса при действии механического стимула на кожу. Вибрацию воспринимают тельца Пачини, располагающиеся как в слизистой, так и на не покрытых волосами частях кожи, в жировой ткани подкожных слоев, а также в суставных сумках, сухожилиях. Эти рецепторы представлены нервными терминалями, заключенными в слоистые оболочки из соединительной ткани. Тельца Пачини обладают очень быстрой адаптацией и реагируют на ускорение при смещении кожи в результате действия механических стимулов, одновременно вовлекаются в реакцию несколько телец Пачини. Щекотание воспринимают свободно лежащие, неинкапсулированные нервные окончания, расположенные в поверхностных слоях кожи. Для данного вида рецепторов характерна низкая специфичность реакции на стимулы разной интенсивности. По функциональным особенностям тактильные рецепторы подразделяются на: - фазные - статические. Фазные тактильные рецепторы возбуждаются при динамическом раздражении. Они обладают высокой чувствительностью, коротким латентным периодом, быстро адаптируются. Статические тактильные рецепторы возбуждаются в основном от статического раздражения. Они менее чувствительны, чем фазные, с более длительным латентным периодом, медленно адаптируются. 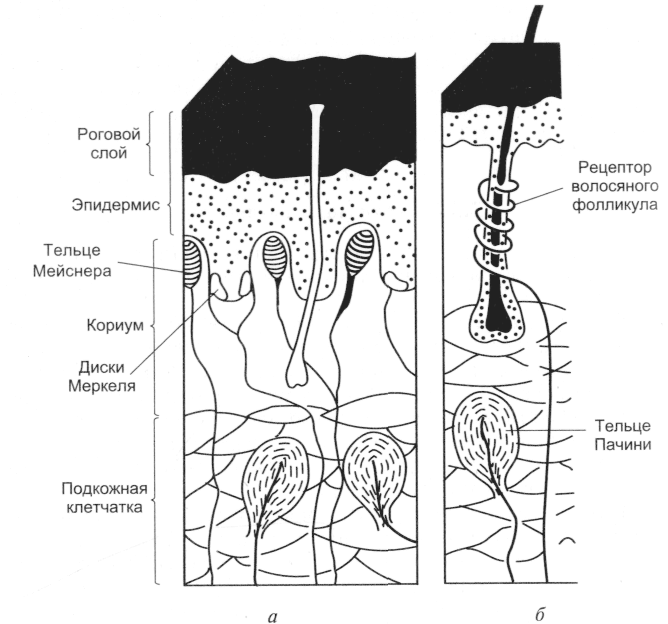 Проводниковый отдел. От большинства механорецепторов в спинной мозг информация поступает в центральную нервную систему по А-волокнам и лишь от рецепторов щекотки – по С-волокнам. Первый нейрон находится в спинальных ганглиях. В заднем роге спинного мозга происходит первое переключение на интернейроны (второй нейрон), от них восходящий путь в составе заднего столба достигает ядер заднего столба в продолговатом мозге (третий нейрон), где происходит второе переключение, далее через медиальную петлю путь следует к вентро-базальным ядрам зрительного бугра (четвертый нейрон), центральные отростки нейронов зрительного бугра идут в кору больших полушарий. Центральный отделтактильного анализатора локализуется в 1 и II зонах соматосенсорной области коры большого мозга (задняя центральная извилина). Строение и функции соматосенсорного анализатора. Двигательная часть. Соматосенсо́рная система — это комплексная система, образованная рецепторами и центрами обработки нервной системы, осуществляющая такие сенсорные модальности, как осязание, температура, проприоцепция, ноцицепция. Соматосенсорная система также осуществляет контроль пространственного положения частей тела между собой. Необходима для выполнения сложных движений, управляемых корой головного мозга. Проявлением деятельности соматосенсорной системы является так называемое «мышечное чувство». Кинесте́зия— так называемое «мышечное чувство», чувство положения и перемещения как отдельных членов, так и всего человеческого тела. Это способность головного мозга постоянно осознавать положение и движение мышц различных частей тела. Эта способность достигается за счёт проприоцепторов, которые посылают в головной мозг импульсы от мышц, суставов и сухожилий. Без такой способности человек не мог бы выполнять координированных движений с закрытыми глазами. Проприоцепция— ощущение своего тела. Ноцице́пция; Физиологи́ческая боль — это активность в афферентных (приносящих) нервных волокнах периферийной и центральной нервной системы, возбуждаемая разнообразными стимулами, обладающими «повреждающей» интенсивностью. Данная активность генерируется ноцицепторами, или по-другому рецепторами боли, которые могут отслеживать механические, тепловые или химические воздействия, превышающие генетически установленный порог обычного восприятия. Получив повреждающий стимул, ноцицептор передаёт сигнал через спинной мозг далее в головной мозг. Роль температурного анализатора в восприятии температуры внешней и внутренней среды организма. Особенности функционирования различных отделов температурного анализатора, его роль в поддержании температурного гомеостаза. Температурные рецепторы имеют важное значение для поддержания постоянной температуры тела. В средней полосе России колебание температуры окружающей среды между различными областями в течение одного дня может достигать 20-25°C, а на протяжении всего года — до 70°C (от 35°С ниже нуля — зимой, до 35°C выше нуля — летом). Без температурной адаптации человек не смог бы выжить. Поэтому очень важны быстрое и точное восприятие изменений температуры и соответствующая перестройка механизмов теплопродукции и теплоотдачи в зависимости от изменившихся условий. Именно в этом состоит функция температурных рецепторов. Полагают, что существуют две разновидности: одни воспринимают тепло, другие — холод. Рецепторы, воспринимающие холод, располагаются ближе к поверхности кожи, их количество больше, чем тепловых, которые и располагаются значительно глубже.Наиболее чувствительна к воздействию температурных раздражителей кожа лица и живота. Кожа ног по сравнению с кожей лица в два раза менее чувствительна к холоду и в четыре — к теплу. Температурные раздражители помогают ощущать структуру комбинации движений и скорость. Происходит это потому, что при быстром изменении положения частей тела или большой скорости передвижения возникает прохладный ветерок. Он воспринимается температурными рецепторами как изменение температуры кожи, а осязательными — как прикосновение воздуха. 5. Терморецепция. Изменение температуры внутренней среды («ядра») и поверхностных отделов («оболочки») тела человека воспринимается организмом с помощью терморецепторов. Температурная рецепция осуществляется окончаниями тонких чувствительных нервных волокон типа С и А (δ), которые представлены в коже, слизистых оболочках, мышцах, сосудах, во внутренних органах (периферические терморецепторы). Холодо- и теплочувствительные нейроны располагаются в медиальной преоптической области переднего гипоталамуса (центральные терморецепторы).   Восприятие температурных раздражений из внешней среды и формирование температурных ощущений у человека осуществляется с помощью терморецепторов кожи и слизистых оболочек, среди которых имеются холодовые рецепторы (повышают частоту передачи нервных импульсов по афферентным нервным волокнам к терморегуляторному центру при их охлаждении и снижают эту частоту при их нагревании) и тепловые рецепторы (реагируют на изменение температуры тела противоположным образом). В коже и на слизистых оболочках человека больше холодовых рецепторов (около 250 000), чем тепловых (около 30 000). Кроме того, холодовые рецепторы кожи расположены более поверхностно, на глубине 0,17 мм, а тепловые — более глубоко, на глубине 0,3 мм. Эта особенность расположения терморецепторов обусловливает более раннее восприятие организмом человека холода, чем тепла. Другая особенность терморецепторов — их неравномерное распределение в коже по площади, что определяет различный уровень чувствительности к холоду и теплу разных участков тела. Наибольшей чувствительностью обладает кожа лица, наименьшей — кожа нижних конечностей. Афферентный поток нервных импульсов от периферических терморецепторов поступает через задние корешки спинного мозга к вставочным нейронам задних рогов. Затем по спиноталамическому тракту этот поток импульсов достигает передних ядер таламуса и далее проводится в соматосенсорную кору больших полушарий головного мозга. Поступление нервных импульсов от периферических терморецепторов в соматосенсорную кору обеспечивает возникновение и топическую локализацию субъективных температурных ощущений, таких как «тепло», «холодно», «прохладно», «жарко», «температурный комфорт» или «дискомфорт». На их основе формируются поведенческие терморегуляторные реакции. Значительная часть афферентных импульсов от периферических рецепторов кожи и внутренних органов поступает из спинного мозга по волокнам спиноталамического тракта к нейронам гипоталамического центра терморегуляции.  6. Проприоцептивная чувствительность Проприорецепторы, проприоцепторы, чувствительные рецепторы (от лат. proprius — «собственный, особенный» и receptor — «принимающий») — периферические элементы сенсорных органов, расположенные в мышцах, связках, суставных сумках, в коже и свидетельствующие об их работе (сокращения мышц, изменения положения тела в пространстве). Проприорецепторы являются частным видом механорецепторов. Чувство, соответствующее этим элементам сенсорных органов, — проприоцепция. К проприоцепторам относят, помимо свободных нервных окончаний, также мышечные веретена, тельца Гольджи, сосредоточенные в сухожилиях, и тельца Пачини, расположенные в фасциях, сухожилиях, связках. Механореце́пторы — это окончания чувствительных нервных волокон, реагирующие на механическое давление или иные механические воздействия: как действующие извне (тактильные рецепторы), так и возникающие во внутренних органах. Типы проприорецепторов: Выделяется 3 типа проприорецепторов. Мышечные веретена (рецепторы растяжения) представляют собой специализированные мышечные волокна, чувствительные к растяжению. Такие волокна окружены соединительнотканной капсулой, вследствие этого их называют интрафузальными. К каждому мышечному веретену подходит отросток спинального чувствительного нейрона (афферентное волокно), которое обвивается вокруг центральной части веретена. Мышечные веретена имеют также и двигательную иннервацию. Сухожильные органы Гольджи (рецепторы мышечной силы) состоят из сухожильных нитей, отходящих примерно от 10 экстрафузальных мышечных волокон и заключенных в соединительнотканную капсулу. К ней подходят 1-2 афферентных нервных волокна. Войдя в капсулу, волокна образуют сильно разветвленные окончания среди сухожильных нитей. Суставные рецепторы располагаются в суставных сумках. Они относятся к механорецепторам, посылающим в ЦНС информацию о положении, деформации и смещении различных частей тела. Функционирование этих рецепторов обеспечивает координацию всех подвижных органов и тканей человека в состоянии покоя и во время любых двигательных актов. Электрические ответы проприорецепторов: Мышечные веретена в ненапряженной мышце генерируют нервные импульсы с некоторой частотой. При растяжении веретена частота нервных импульсов увеличивается, при уменьшении длины веретена (например, при сокращении мышцы)-уменьшается. Таким образом мышечные веретена воспринимают длину мышцы. Сухожильные органы в состоянии покоя «молчат», они начинают генерировать импульсы при натяжении сухожилий, например, при сокращении мышцы или при ее пассивном растяжении под действием груза. Таким образом сухожильные органы воспринимают натяжение мышцы. Суставные рецепторы активируются при изменении положения сустава. Механизмы проприорецепии: Проприорецепторы обеспечивают наличие следующих видов чувствительности: Чувство позы – даже с закрытыми глазами возможно почувствовать, под какими углами располагаются суставы нашего тела; Чувство движения – при изменении положения сустава ощущается направление и скорость этого движения; Чувство силы позволяет оценить интенсивность совершаемого мышечного усилия; это же чувство лежит в основе определения веса предметов. Информация от проприорецепторов поступает в соматосенсорную кору, главным образом, по системе задних столбов. На уровне таламуса и коры происходит интегрирование информации от проприорецепторов, механорецепторов кожи, зрительной системы, вестибулярной системы. В результате этого строится интегральная модель тела в трехмерном пространстве. 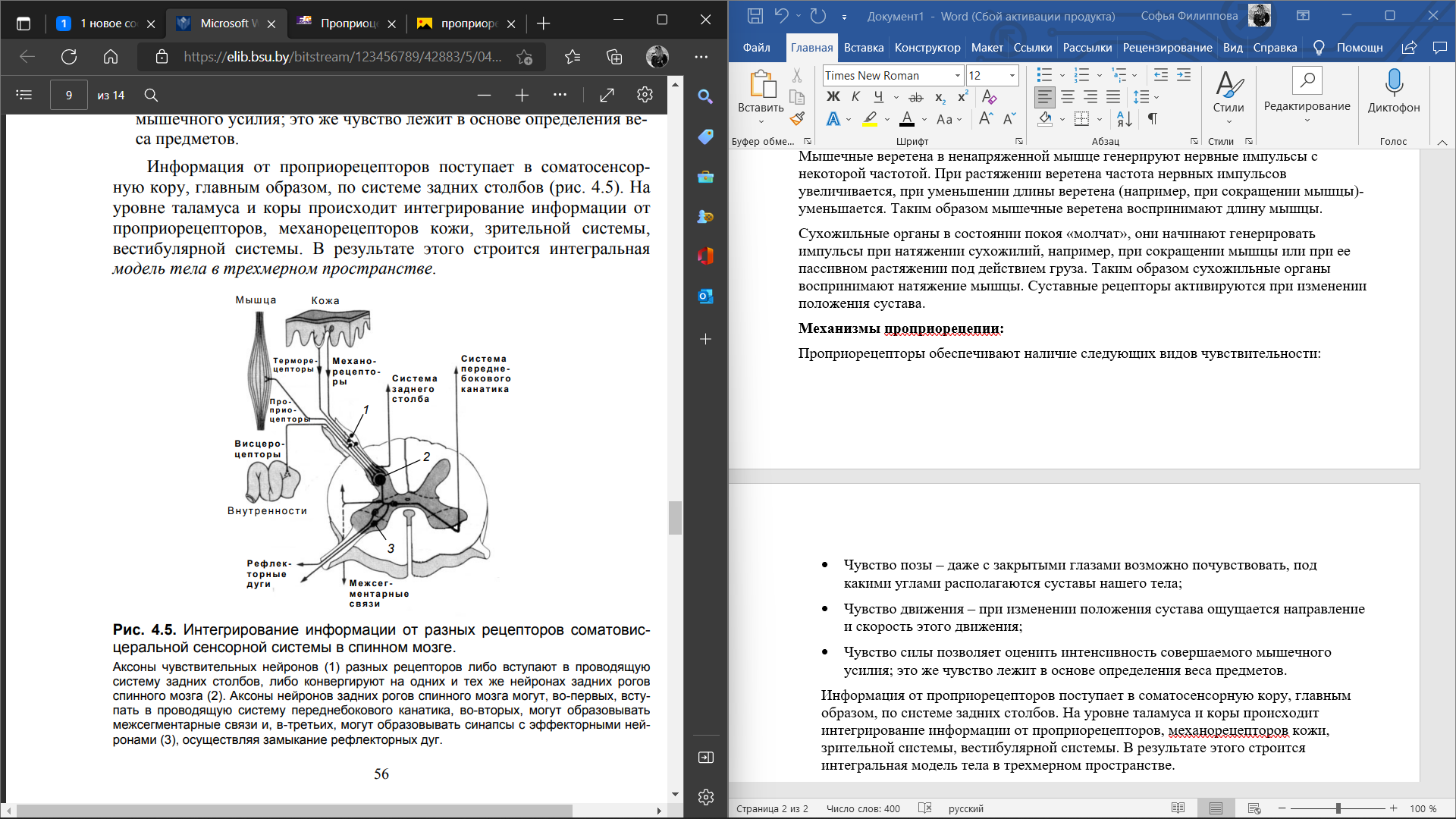 7. Висцеральная чувствительность.  Висцеральная чувствительность Существует несколько типов висцерорецепторов (интерорецепторов): • барорецепторы воспринимают давление биологических жидкостей (например, баорорецепторы каротидного синуса); • рецепторы растяжения воспринимают степень растяжения органов (например, полых вен, легких, мочевого пузыря); • хеморецепторы чувствительны к изменениям концентрации определенных веществ в жидкостях внутренней среды организма (О2- и СО2-рецепторы, рецепторы рН, глюкорецепторы и др.); • осморецепторы активируются при изменении осмотического давления крови, межклеточной жидкости. Информация от висцерорецепторов используется главным образом для процессов регуляции жизнедеятельности. Эта информация по переднебоковой системе достигает коры больших полушарий и частично осознается. Связь висцерорецепторов с проводящими путями соматосенсорной системы показана на рис. 4.5. Вместе с тем, лишь немногие изменения во внутренней среде организма могут быть сознательно восприняты. Например, снижение парциального давления О2 не ощущается, даже если оно достаточно сильное, чтобы вызвать полубессознательное состояние. Активация рецепторов растяжения желудка вызывает чувство наполнения и сытости, а возбуждение осморецепторов может вызывать чувство жажды. Раздражение рецепторов почек и мочеточников всегда воспринимается как боль. Растяжение стенки мочевого пузыря обусловливает потребность мочеиспускания. 8. Зрительная сенсорная система и её роль в регуляции движений. Зрительная система передаёт мозгу более 90% сенсорной информации. Зрение – многозвеньевой процесс, начинающийся с проекции изображения на сетчатке глаза, затем происходит возбуждение фоторецепторов, передача и преобразование зрительной информации в нейронных слоях зрительной системы. Заканчивается зрительное восприятие формированием в затылочной доле коры больших полушарий зрительного образа. Периферический отдел зрительного анализатора представлен органом зрения (глазом), который служит для восприятия световых раздражений и находится в глазнице. Орган зрения состоит из глазного яблока и вспомогательного аппарата. Проводниковый отдел зрительного анализатора начинается зрительным нервом, который направляется из глазницы в полость черепа. В полости черепа зрительные нервы образуют частичный перекрёст, причём, нервные волокна, идущие от наружных (височных) половинок сетчатки, не перекрещиваются, оставаясь на своей стороне, а волокна, идущие от внутренних (носовых) половин её, перекрещиваясь, переходят на другую сторону. Различие функций палочек и колбочек и механизм фоторецепции. Целый ряд факторов свидетельствует о то, что палочки являются аппаратом сумеречного зрения, т. е. функционируют в сумерках, а колбочки – аппаратом дневного зрения. Колбочки воспринимают лучи в условиях яркой освещённости. С их деятельностью связано восприятие цвета. О различиях в функциях палочек и колбочек свидетельствует структура сетчатки разных животных. Так, сетчатка дневных животных – голубей, ящериц и др. – содержит преимущественно колбочки, а ночных (например, летучих мышей) – палочки. Наиболее отчётливо воспринимается цвет при действии лучей на область центральной ямки, если же они попадают на периферию сетчатки, то возникает бесцветное изображение. При действии лучей света на наружном сегменте палочек зрительный пигмент родопсин разлагается на ретиналь – производное витамина А и белок опсин. На свету после отделения опсина происходит превращение ретиналя напосредственно в витамин А, который из наружных сегментов перемещается в клетки пигментного слоя. Считают, что витамин А увеличивает проницаемость клеточных мембран. В темноте происходит восстановление родопсина, для чего необходим витамин А. При его недостатке возникает нарушение видения в темноте, что называют куриной слепотой. В колбочках имеется светочувствительное вещество, сходное с родопсином, его называют йодопсином. Оно тоже состоит из ретиналя и белка опсина, но структура последнего неодинакова с белком родопсина. Вследствие целого ряда химических реакций, которые протекают в фоторецепторах, в отростках ганглиозных клеток сетчатки возникает распространяющееся возбуждение, направляющееся в зрительные центры головного мозга. Зрительная адаптация. При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной сенсорной системы к условиям яркой освещённости называется световой адаптацией. Обратное явление (темновая адаптация) наблюдается при переходе из светлого помещения в почти неосвещённое. В первое время человек почти ничего не видит из-за пониженной возбудимости фоторецепторов и зрительных нейронов. Постепенно начинают выявляться контуры предметов, а затем различаются и их детали, так как чувствительность фоторецепторов и зрительных нейронов в темноте постепенно повышается. Повышение световой чувствительности во время пребывания в темноте происходит неравномерно: в первые 10 минут она увеличивается в десятки раз, а затем в течение часа – в десятки тысяч раз. Важную роль в этом процессе играет восстановление зрительных пигментов. Пигменты колбочек в темноте восстанавливаются быстрее родопсина палочек, поэтому в первые минуты пребывания в темноте адаптация обусловлена процессами в колбочках. Этот первый период адаптации не приводит к большим изменениям чувствительности глаза, так как абсолютная чувствительность колбочкового аппарата невелика. Следующий период адаптации обусловлен восстановлением родопсина палочек. Этот период завершается только к концу первого часа пребывания в темноте. Восстановление родопсина сопровождается резким (в 100000 – 200000 раз) повышением чувствительности палочек к свету. В связи с максимальной чувствительностью в темноте только палочек, слабо освещённый предмет виден лишь периферическим зрением. Роль движения глаз для зрения. При рассматривании любых предметов глаза двигаются. Глазные движения осуществляют 6 мышц, прикреплённых к глазному яблоку. Движения двух глаз совершаются одновременно и содружественно. Рассматривая близкие предметы, необходимо сводить, а рассматривая далёкие предметы – разводить зрительные оси двух глаз. Важная роль движений глаз для зрения определяется также тем, что для непрерывного получения мозгом зрительной информации необходимо движение изображения на сетчатке. Импульсы в зрительном нерве возникают в момент включения и выключения светового изображения. При длящемся действии света на одни и те же фоторецепторы импульсация в волокнах зрительного нерва быстро прекращается и зрительное ощущение при неподвижных глазах и объектах исчезает через 1-2 с. Чтобы этого не случилось, глаз при рассматривании любого предмета производит не ощущаемые человеком непрерывные скачки. Вследствие каждого скачка изображение на сетчатке смещается с одних фоторецепторов на новые, вновь вызывая импульсацию ганглиозных клеток. Продолжительность каждого скачка равна сотым долям секунды, а амплитуда его не превышает 20º. Чем сложнее рассматриваемый объект, тем сложнее траектория движения глаз. Они как бы прослеживают контуры изображения, задерживаясь на наиболее информативных его участках (например, в лице – это глаза). Кроме того, глаз непрерывно мелко дрожит и дрейфует (медленно смещается с точки фиксации взора) – саккады. Эти движения также играют роль в дезадаптации зрительных нейронов. 9. Сетчатка. Сетчатка — внутренняя оболочка глаза являющаяся периферическим, начальным звеном зрительного анализатора, который обеспечивает восприятие и преобразование видимой части спектра в нервные импульсы. - светочувствительная часть глаза, которая содержит: колбочки, ответственные за цветовое зрение; палочки, ответственные в основном за черно-белое зрение и зрение в темноте. Сетчатку можно характеризовать как часть мозга, вынесенную на периферию. Сетчатка выстилает внутреннюю поверхность глазного яблока. Она плотно прикреплена только у зубчатой линии и около диска зрительного нерва. Остальная (большая) ее часть прилежит к сосудистой оболочке свободно и удерживается только давлением стекловидного тела и тонкими связями пигментного эпителия, что объясняет возможность развития отслойки сетчатки. При возбуждении палочек и колбочек сигналы сначала проводятся через последовательные слои нейронов самой сетчатки, затем — в нервные волокна зрительных путей и в итоге — в кору большого мозга. Целью этой главы является объяснение механизмов, с помощью которых палочки и колбочки воспринимают свет и цвет и преобразуют зрительный образ в сигналы зрительного нерва. Анатомия и функция структурных элементов сетчатки. Слои сетчатки: Первый слой – пигментный эпителий сетчатки. Он прилежит к мембране Бруха хориоидеи. Второй слой – наружные сегменты палочек и колбочек (нейронов первого порядка). Здесь происходят первичные фотофизические и ферментативные процессы трансформации энергии света в физиологическое возбуждение. Третий слой – наружная пограничная мембрана. Это полоса межклеточных сцеплений. Четвертый слой – наружный ядерный. Образован телами и ядрами фоторецепторов – палочек и колбочек (нейронов первого порядка). Пятый слой – наружный плексиформный,или сетчатый(от лат. plexus – сплетение), соединение нейронов первого и второго порядка. Шестой слой – внутренний ядерный. Его образуют ядра биполярных клеток (нейронов второго порядка), а также ядра амакриновых, горизонтальных и мюллеровских клеток, обеспечивающих скелетную, ассоциативную и трофическую функцию. Седьмой слой – внутренний плексиформный – синаптическая зона нейронов второго и третьего порядка. Является границей между наружной бессосудистой частью сетчатки и внутренней, в которой располагаются собственные сосуды сетчатки. Питание наружного бессосудистого слоя сетчатки обеспечивает хориоидея. Восьмой слой образован ганглиозными клетками (нейронами третьего порядка). Девятый слой состоит из аксонов ганглиозных клеток, они сходятся в общий ствол в диске зрительного нерва. Десятый слой – внутренняя пограничная мембрана,отделяющая сетчатку от пограничной мембраны стекловидного тела. После того как свет проходит через оптическую систему глаза и стекловидное тело, он входит в сетчатку изнутри (для облегчения понимания просим вас изучить рисунок выше). Прежде чем свет достигнет слоя палочек и колбочек, расположенного по всему наружному краю глаза, он проходит через ганглиозные клетки, сетчатые и ядерные слои. Толщина преодолеваемого светом слоя составляет несколько сотен микрометров, и этот путь через негомогенную ткань снижает остроту зрения. Однако в области центральной ямки сетчатки внутренние слои раздвинуты в стороны для уменьшения этой потери зрения. Область центральной ямки сетчатки Ямка-небольшая область в центре сетчатки, общей площадью немногим более 1 мм; она имеет особую способность к острому детальному видению. Эта центральная ямка диаметром лишь 0,3 мм почти полностью состоит из колбочек, особая структура которых помогает выявлять детали в зрительном образе. В частности, у колбочек центральной ямки особо длинные и тонкие тела в отличие от гораздо более толстых колбочек на периферии сетчатки. Кроме того, в области ямки кровеносные сосуды, ганглиозные клетки, внутренний ядерный слой клеток и сетчатые слои лежат не над колбочками, а смещены в сторону. Это позволяет свету проходить к колбочкам беспрепятственно. Палочки и колбочки Наружный сегмент колбочки имеет форму конуса. В целом палочки уже и длиннее колбочек, но не всегда. Так, в периферических частях сетчатки палочки имеют диаметр 2-5 мкм, а колбочки — 5-8 мкм; в центральной ямке колбочки тоньше и имеют диаметр лишь 1,5 мкм. Главные функциональные сегменты фоторецепторов: (1) наружный сегмент; (2) внутренний сегмент; (3) ядро; (4) синаптическое тело. Светочувствительное вещество находится в наружном сегменте; для палочек это родопсин, а в колбочках обнаруживается одно из трех «цветных» фоточувствительных веществ, обычно называемых просто цветными пигментами и функционирующих практически так же, как родопсин, кроме различий в спектральной чувствительности. В наружных сегментах палочек и колбочек большое количество дисков. Они фактически представляют собой складки клеточной мембраны, «упакованные» в стопку. В каждой палочке или колбочке содержится примерно по 1000 дисков. И родопсин, и цветные пигменты — конъюгированные белки. Они включены в мембраны дисков в виде трансмембранных белков. Концентрация этих фоточувствительных пигментов в дисках так велика, что на их долю приходится около 40% всей массы наружного сегмента. Внутренний сегмент палочки или колбочки содержит обычную цитоплазму с цитоплазматическими органеллами. Особое значение имеют митохондрии — они играют важную роль в обеспечении фоторецепторной функции энергией. Синаптическое тело — часть палочек и колбочек, которая соединяется с последующими нервными клетками (горизонтальными и биполярными), представляющими следующие звенья зрительного пути. 10. Слуховая сенсорная система и её роль в регуляции движений. Функция слуховой системы состоит в формировании слуховых ощущений человека в ответ на действие звуковых волн, представляющих собой распространяющиеся колебания молекул воздуха (упругой среды). Периферическая часть слуховой системы включает наружное, среднее и внутреннее ухо, в котором расположены слуховые рецепторы. Ее центральную часть образуют проводящие пути, переключательные ядра и слуховая кора, расположенная в обоих полушариях в глубине латеральной борозды, отделяющей височную долю от лобной и передних отделов теменной доли (поля 41 и 42). Адекватный раздражитель – звуки – колебательные движения частиц упругих тел, распределяющиеся в различных средах в виде волн. Звуковые волны характеризуются частотой и амплитудой. Частота определяет высоту звука. Второй характеристикой звука является сила или интенсивность (громкость) звука, которая зависит от амплитуды звуковых колебаний. Единицей измерения громкости является Бел, хотя в практике используется децибел (0,1 Бел). Чувствительность слухового анализатора определяется минимальной силой звука, достаточной для возникновения слухового ощущения. Звуковые волны представляют собой передаваемые от источника звука механические смещения молекул воздуха (или иной упругой среды). Скорость распространения звуковых волн в воздухе около 343 м/с при 20 °С (в воде и металлах она значительно выше). Правильно чередующиеся участки сжатия и разрежения молекул упругой среды можно представить в виде синусоид, которые различаются частотой и амплитудой. При суперпозиции звуковых волн с различными частотами и амплитудами они наслаиваются друг на друга, образуя комплексные волны. Физическим понятиям амплитуды, частоты и комплексности соответствуют ощущения громкости, высоты и тембра звука. 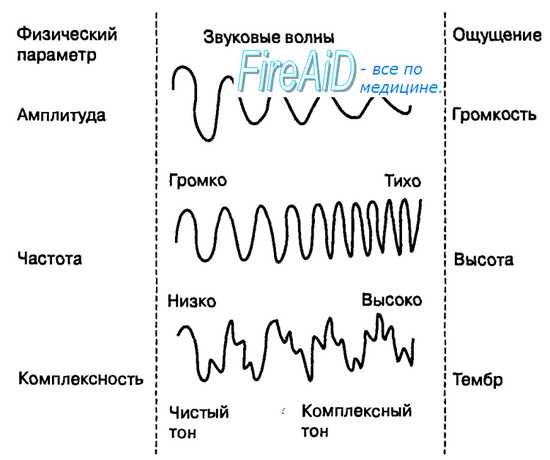 Звук, образованный синусоидальными колебаниями только одной частоты, вызывает ощущение определенной его высоты и определяется как тон. Комплексные тоны состоят из основного тона (самая низкая частота колебаний) и определяющих тембр обертонов, или гармоник, представляющих более высокие частоты, кратные основной. В повседневной жизни тоны всегда бывают комплексными, т. е. составленными из нескольких синусоид. Индивидуальное сочетание комплексных волн определяет характерный тембр человеческого голоса или музыкального инструмента. Слуховая система человека способна различать высоту звука лишь у периодических звуковых сигналов, тогда как звуковые раздражители, состоящие из беспорядочного сочетания частотных и амплитудных компонентов, воспринимаются как шум. Показатели слуха: Диапазон слышимых частот (15 (20) -20 тыс. Гц). Ниже 20 Гц – инфразвуки, выше 20тыс. Гц – ультразвуки (человек не ощущает). Наибольшая чувствительность наблюдается в области звуковых колебаний от 1 тыс. – 3 тыс. в секунду, соответствующей человеческой речи (речевому диапазону). Дифференциальная чувствительность. Способность различать минимальные изменения параметров звука. В области средних частот от 500 – 2 тыс. Гц. Дифференциальный порог по интенсивности составляет 0,5 – 1 дБел по частоте 1%. Пространственная и временная разрешающая способность. 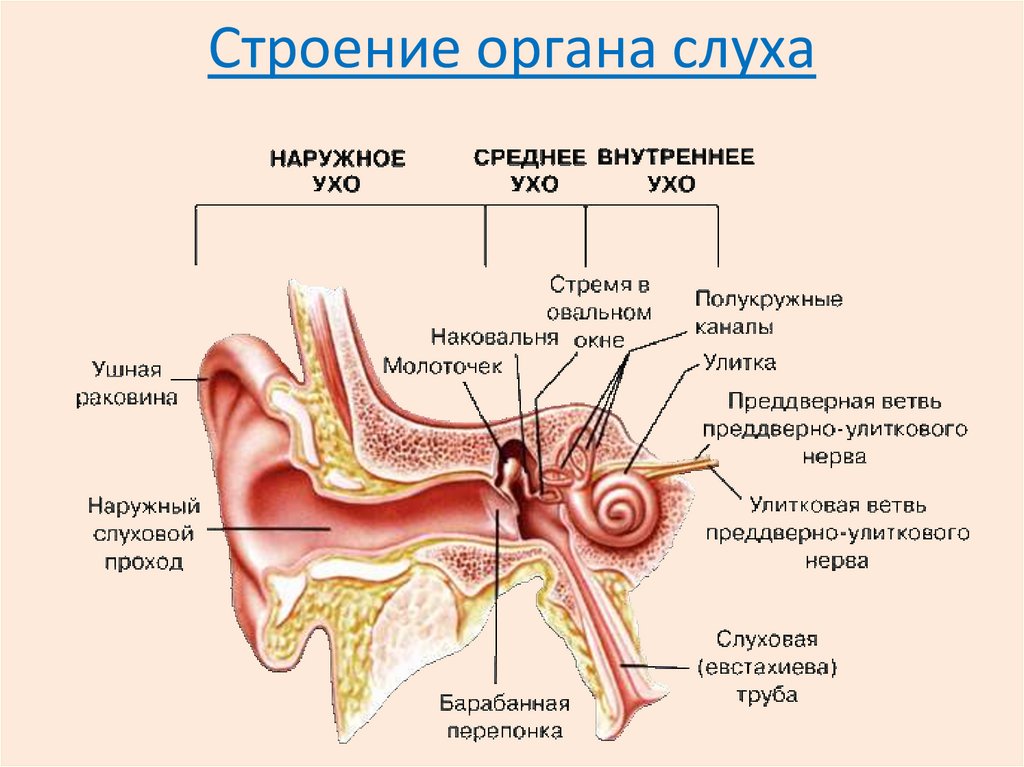 Пространственная локализация звука обеспечивается преимущественно за счет двух разных механизмов. Их включение зависит от частоты звука или его длины волны. При низкочастотных сигналах (примерно до 1,5 кГц) длина волны оказывается меньше межушного расстояния, равного в среднем у человека 21 см. В этом случае локализация источника осуществляется благодаря разному времени прихода звуковой волны на каждое ухо в зависимости от азимута. При частотах больше 3 кГц длина волны заведомо меньше межушного расстояния. Такие волны не могут обогнуть голову, они многократно отражаются от окружающих предметов и головы, теряя при этом энергию звуковых колебаний. В этом случае локализация осуществляется в основном за счет межушных различий по интенсивности. В области частот от 1,5 Гц до 3 кГц происходит смена временного механизма локализации на механизм оценки интенсивности, а область перехода оказывается неблагоприятной для определения местонахождения источника звука. При определении местонахождения источника звука важно оценить его удаленность. Существенную роль в решении этой задачи играет интенсивность сигнала: чем больше расстояние от наблюдателя, тем меньше воспринимаемая интенсивность. При больших расстояниях (более 15 м) мы учитываем спектральный состав дошедшего до нас звука: звуки высокой частоты затухают быстрее, т.е. «пробегают» меньшее расстояние, звуки низкой частоты, напротив, затухают медленнее и распространяются дальше. Именно поэтому звуки, издаваемые удаленным источником, кажутся нам более низкими. Одним из факторов, существенно облегчающих оценку удаленности, является реверберация звукового сигнала от отражающих поверхностей, т.е. восприятие отраженного звука. Слуховая система способна определять не только местоположение неподвижного, но и движущегося источника звука. Физиологической основой оценки локализации источника звука является активность так называемых нейронов-детекторов движения, расположенных в верхнеоливарном комплексе, задних холмах, внутреннем коленчатом теле и слуховой коре. Но ведущая роль здесь принадлежит верхним оливам и задним холмам. 11. Проведение звука. Существует два пути проведения звуков: воздушная проводимость: через наружный слуховой проход, барабанную перепонку и цепь слуховых косточек; тканевая проводимость: через ткани черепа. Функция слухового анализатора: восприятие и анализ звуковых раздражений. Периферический отдел: слуховые рецепторы в полости внутреннего уха. Проводниковый отдел: слуховой нерв. Центральный отдел: слуховая зона в височной доле коры больших полушарий. Строение уха Орган слуха у человека расположен в полости черепа в толще височной кости. Он делится на три отдела: наружное, среднее и внутреннее ухо. Эти отделы тесно связаны анатомически и функционально. Наружное ухо состоит из наружного слухового прохода и ушной раковины. Среднее ухо — барабанная полость; она отделена барабанной перепонкой от наружного уха. Внутреннее ухо, или лабиринт, — отдел уха, где происходит раздражение рецепторов слухового (улиткового) нерва; он помещается внутри пирамиды височной кости. Внутреннее ухо образует орган слуха и равновесия. Наружное и среднее ухо имеют второстепенное значение: они проводят звуковые колебания к внутреннему уху, и таким образом является звукопроводящим аппаратом. Наружное ухоНаружное ухо включает ушную раковину и наружный слуховой проход, которые предназначены для улавливания и проведения звуковых колебаний. Ушная раковина образована тремя тканями: тонкой пластинкой гиалинового хряща, покрытого с обеих сторон надхрящницей, имеющего сложную выпукло-вогнутую форму, определяющую рельеф ушной раковины; кожей очень тонкой, плотно прилегающей к надхрящнице и почти не имеющей жировой клетчатки; подкожной жировой клетчаткой, расположенной в значительном количестве в нижнем отделе ушной раковины — мочке уха. Ушная раковина прикрепляется к височной кости связками и имеет рудиментарные мышцы, которые хорошо выражены у животных. Ушная раковина устроена так, чтобы максимально концентрировать звуковые колебания и направлять их в наружное слуховое отверстие. Форма, величина, постановка ушной раковины и размеры ушной дольки индивидуальны у каждого человека. Дарвинов бугорок — рудиментарный треугольный выступ, который наблюдается у 10% людей в верхне-задней области завитка раковины; он соответствует верхушке уха животных. Наружный слуховой проход представляет собой S-образную трубку длинной примерно 3 см и диаметром 0,7 см, которая снаружи открывается слуховым отверстием и отделяется от полости среднего уха барабанной перепонкой. Хрящевая часть, являющаяся продолжением хряща ушной раковины, составляет 1/3 его длины, остальные 2/3 образованы костным каналом височной кости. В месте перехода хрящевого отдела в костный канал сужается и изгибается. В этом месте находится связка из эластичной соединительной ткани. Такое строение делает возможным растяжение хрящевого отдела прохода в длину и в ширину. В хрящевой части слухового прохода кожа покрыта короткими волосками, предохраняющими от попадания в ухо мелких частиц. В волосяные фолликулы открываются сальные железы. Характерным для кожи этого отдела является наличие в более глубоких слоях серных желез. Серные железы являются производными потовых желез.Серные железы впадают либо в волосяные фолликулы, либо свободно в кожу. Серные железы выделяют светло-желтый секрет, который вместе с отделяемым сальных желез и с отторгшимся эпителием образует ушную серу. Ушная сера — светло-желтый секрет серных желез наружного слухового прохода. Сера состоит из белков, жиров, жирных кислот и минеральных солей. Часть белков являются иммуноглобулинами, определяющими защитную функцию. Кроме того, в состав серы входят отмершие клетки, кожное сало, пыль и другие включения. Функция ушной серы: увлажнение кожи наружного слухового прохода; очистки слухового прохода от инородных частиц (пыли, сора, насекомых); защита от бактерий, грибков и вирусов; жировая смазка в наружной части слухового прохода препятствует попаданию в него воды. Ушная сера вместе с загрязнениями естественным образом выводится из слухового прохода наружу при жевательных движениях и речи. Кроме этого кожа слухового прохода постоянно обновляется и растет наружу из слухового прохода, вынося с собой серу. Внутренний костный отдел наружного слухового прохода является каналом височной кости, заканчивающимся барабанной перепонкой. В середине костного отдела расположено сужение слухового прохода — перешеек, за которым расположен более широкий участок. Кожа костного отдела тонкая, не содержит волосяных луковиц и желез и переходит на барабанную перепонку, образуя ее наружный слой. Барабанная перепонка представляет собой тонкую овальную (11 x 9 мм) полупрозрачную пластинку, непроницаемую для воды и воздуха. Перепонка состоит из эластических и коллагеновых волокон, которые в верхней ее части замещены волокнами рыхлой соединительной ткани. Со стороны слухового прохода перепонка покрыта плоским эпителием, а со стороны барабанной полости — эпителием слизистой оболочки. В центральной части барабанная перепонка вогнута, к ней со стороны барабанной полости прикрепляется рукоятка молоточка — первой слуховой косточки среднего уха. Среднее ухоСреднее ухо включает выстланную слизистой оболочкой и заполненную воздухом барабанную полость (объем около 1 см), три слуховые косточки и слуховую (евстахиеву) трубу. Барабанная полость находится в толщине височной кости, между барабанной перепонкой и костным лабиринтом. В барабанной полости помещаются слуховые косточки, мышцы, связки, сосуды и нервы. Стенки полости и все органы, находящиеся в ней, покрыты слизистой оболочкой. В перегородке, отделяющей барабанную полость от внутреннего уха, находится два окна: овальное окно: находится в верхней части перегородки, ведет в преддверие внутреннего уха; закрыто основанием стремечка; круглое окно: расположено в нижней части перегородки, ведет в начало улитки; закрыто вторичной барабанной перепонкой. В барабанной полости находятся три слуховые косточки: молоточек, наковальня и стремя (= стремечко). Слуховые косточки имеют небольшие размеры. Соединяясь между собой, они образуют цепь, которая тянется от барабанной перепонки до овального отверстия. Все косточки соединяются между собой при помощи суставов и покрыты слизистой оболочкой. Молоточек рукояткой сращен с барабанной перепонкой, а головкой при помощи сустава соединяется с наковальней, которая в свою очередь подвижно соединена со стременем. Основание стремени закрывает овальное окно преддверия. Мышцы барабанной полости (натягивающая барабанную перепонку и стременная) удерживают слуховые косточки в состоянии напряжения и защищают внутреннее ухо от чрезмерных звуковых раздражений. Слуховая (евстахиева) труба соединяет барабанную полость среднего уха с носоглоткой. Это мышечная трубка, которая раскрывается при глотании и зевании. Слизистая оболочка, выстилающая слуховую трубу, является продолжением слизистой оболочки носоглотки, состоит из мерцательного эпителия с движением ресничек из барабанной полости в носоглотку. Функции евстахиевой трубы: уравновешивание давления между барабанной полостью и внешней средой для поддержания нормальной работы звукопроводящего аппарата; защита от проникновения инфекций; удаление из барабанной полости случайно проникших частиц. Внутреннее ухоВнутреннее ухо состоит из костного и вставленного в него перепончатого лабиринта. Костный лабиринт состоит из трех отделов: преддверия, улитки и трех полукружных каналов. Преддверие — полость небольших размеров и неправильной формы, на наружной стенке которого расположены два окна (круглое и овальное), ведущие в барабанную полость. Передняя часть преддверия сообщается с улиткой через лестницу преддверия. Задняя часть содержит два вдавления для мешочков вестибулярного аппарата. Улитка — костный спиральный канал в 2,5 оборота. Ось улитки лежит горизонтально и называется костным стержнем улитки. Вокруг стержня обвивается костная спиральная пластинка, которая частично перегораживает спиральный канал улитки и делит его на лестницу преддверия и барабанную лестницу. Между собой они сообщаются только через отверстие, находящееся у верхушки улитки. Полукружные каналы — костные образования, расположенные в трех взаимно перпендикулярных плоскостях. Каждый канал имеет расширенную ножку (ампулу). Перепончатый лабиринт заполнен эндолимфой и состоит из трех отделов: перепончатой улитки, или улиткового протока, продолжение спиральной пластинки между лестницей предверия и барабанной лестницей. В улитковом протоке находится слуховые рецепторы — спиральный, или кортиев, орган; трех полукружных каналов и двух мешочков, расположенных в преддверии, которые играют роль вестибулярного аппарата. Между костным и перепончатым лабиринтом находится перилимфа --видоизмененная спинномозговая жидкость. |