акустическая коммуникация рыб. 1. Определение понятия Акустическая коммуникация рыб
 Скачать 180.5 Kb. Скачать 180.5 Kb.
|

|
2 Содержание Введение 3 1. Определение понятия «Акустическая коммуникация рыб» 4 2. Способы генерации звуков. 8 3. О чем говорят рыбы ?. 17 Заключение 18 ВведениеРыбы. Утверждение «нем как рыба», давным-давно опровергнуто учеными. Рыбы производят множество звуков, стуча жаберными крышками, и при помощи плавательного пузыря. Каждый вид издает особые звуки. Так, например, морской петух «кудахчет» и «квохчет», ставрида «лает», рыба-барабанщик из породы горбылевых издает шумные звуки, действительно напоминающие барабанный бой, а морской налим выразительно урчит и «хрюкает». Сила звука некоторых морских рыб так велика, что они вызывали взрывы акустических мин, получивших распространение во второй мировой войне и предназначенных, естественно, для поражения кораблей противника. Звуковые сигналы используются для сбора в стаю, как приглашение к размножению, для защиты территории, а также как способ индивидуального распознавания. У рыб нет барабанных перепонок, и они слышат не так, как люди. Система тонких косточек, т.н. веберов аппарат, передает колебания от плавательного пузыря к внутреннему уху. Диапазон частот, которые воспринимают рыбы, сравнительно узок – большинство не слышит звуков выше верхнего «до» и лучше всего воспринимает звуки ниже «ля» третьей октавы. О том, что рыбы способны издавать звуки, было известно еще Аристотелю и Плинию. Однако серьезное изучение этих явлений смогло начаться лишь в 1940-е годы, когда началось конструирование специальной аппаратуры для обнаружения и записи подводных звуков. Основные сведения по этому вопросу, накопленные за последующие 20 лет по темам генерации и восприятия звуков рыбами, были сведены воедино в работе энтузиаста по акустике этих животных У. Таволги[1]. Другой исследователь, Г. Винн, в том же 1964 году обобщил данные по использованию звуков рыбами в разных поведенческих контекстах (звуки, генерируемые спонтанно и в ответ на взятие особи в руки, в конфликтах на основе конкуренции за корм, в момент образования косяков, в нерестовых скоплениях и при их миграциях). Среди прочих рассматривались и такие звуки, в отношении которых предполагалось их использование во внутривидовой коммуникации — при территориальных конфликтах и в половых взаимодействиях самцов и самок. В общей сложности автор использовал данные по 22 видам из 20 родов. Много позже, в 1999 году, в другой обобщающей работе, где внимание было сконцентрировано на роли звуков в акустической коммуникации рыб, в том же аспекте были рассмотрены данные по 25 видам из 15 семейств. Это были представители 19 родов, из которых 15 оказались новыми по сравнению с упомянутыми в статье Винна. Автор этой статьи подчеркивает фрагментарность сведений, которыми мы располагаем по акустическому поведению и их роли в коммуникативном поведении у представителей этого самого обширного класса позвоночных. Автор пишет: «Например, из 69 видов семейства Batrachoididae только для четырех описаны издаваемые ими звуки. Из 209 видов семейства Ophidiidae то же можно сказать лишь в отношении одного вида. Из 285 видов семейства Macrouridae ни один не изучен в этом аспекте». К сказанному следует добавить, что класс рыб объединяет почти 390 семейств. Эти цифры хорошо иллюстрируют меру нашего незнания темы, о которой речь пойдет далее. Особенно неясно, сколь широко акустическая сигнализация представлена у более чем 20 тыс. видов рыб и насколько существенную роль она играет в их жизни. Рыбы используют по крайней мере три типа коммуникативных сигналов: звуковые, зрительные и химические, часто их комбинируя. Рыбы производят звуки, стуча жаберными крышками, а при помощи плавательного пузыря издают ворчание и посвистывание. звуковые сигналы используются для сбора в стаю, как приглашение к размножению, для защиты территории, а также как способ распознавания. у рыб нет барабанных перепонок, и они слышат не так, как люди. система тонких косточек, т.н. веберов аппарат, передает колебания от плавательного пузыря к внутреннему уху. диапазон частот, которые воспринимают рыбы, сравнительно узок – большинство не слышит звуков выше верхнего «до» и лучше всего воспринимает звуки ниже «ля» третьей октавы. 1 Рыбы обладают хорошим зрением, но плохо видят в темноте, например в глубинах океана. Большинство рыб в той или иной степени воспринимает цвет,- это важно в брачный период, поскольку яркая окраска особей одного пола, обычно самцов, привлекает особей противоположного пола. Изменения окраски служат предупреждением для других рыб, говорящим о том, что не следует вторгаться на чужую территорию. В период размножения некоторые рыбы, например трехиглая колюшка, устраивают брачные танцы; другие, например сомики-кошки, демонстрируют угрозу, поворачиваясь широко открытым ртом в сторону чужака. Рыбы, подобно насекомым и некоторым другим животным, используют феромоны – химические сигнальные вещества. Сомики-кошки опознают особей своего вида, ощущая вкус выделяемых ими веществ, вероятно продуцируемых гонадами или содержащихся в моче или слизистых клетках кожи, вкусовые почки сомиков расположены в коже, и любой из них может запомнить вкус феромонов другого, если они хоть раз находились поблизости друг от друга. следующая встреча этих рыб может окончиться войной или миром в зависимости от сложившихся ранее отношений. Принято полагать, что у рыб, как и у членистоногих, способность воспринимать возмущения волнового характера во внешней среде (что кратко и условно обозначается в качестве «слуха») существовала ранее, чем возможность генерации звуков. В самом деле, органы слуха присутствовали уже у наиболее примитивных ископаемых водных форм (так называемых «бесчелюстных»)[2], которых можно считать предковыми по отношению к рыбам как таковым, вымершим и современным. К бесчелюстным относятся ныне живущие миноги и миксины, обладающие внутренним ухом, хотя и более просто устроенным, чем у рыб. Принято полагать, что у рыб, как и у членистоногих, способность воспринимать возмущения волнового характера во внешней среде (что кратко и условно обозначается в качестве «слуха») существовала ранее, чем возможность генерации звуков. В самом деле, органы слуха присутствовали уже у наиболее примитивных ископаемых водных форм (так называемых «бесчелюстных»)[2], которых можно считать предковыми по отношению к рыбам как таковым, вымершим и современным. К бесчелюстным относятся ныне живущие миноги и миксины, обладающие внутренним ухом, хотя и более просто устроенным, чем у рыб. Предполагается, что изначально внутреннее ухо этих существ было органом, контролирующим положение головы и тела относительно направления силы тяжести. В дальнейшем эта структура обрела функцию распознавания низкочастотных колебаний и реагирования на них. Верхняя граница этих частот составляла предположительно около 500 Гц, а на последующих этапах эволюции шел процесс расширения спектра восприятия в более высокочастотные зоны. Восприятие колебаний, поступающих из внешней среды. Ткани тела рыб имеют примерно такую же плотность, как сама вода, в которой они живут. Поэтому колебания ее частиц, чтобы быть воспринятыми организмом, должны встретить на своем пути какую-то структуру внутри тела, которая обладала бы иной плотностью и стала бы сама колебаться в ином ритме в ответ на эти возмущения извне. Таких структур в теле рыбы две. Это, во-первых, три костных образования, именуемых отолитами, которые помещаются в улитке внутреннего уха. В силу того, что их плотность существенно выше, чем у всех прочих тканей, они в ответ на внешние колебания движутся с замедленной скоростью. Эти колебания воспринимаются чувствительными волосковыми клетками, локализованными на стенках каналов улитки, и передаются по системе нейронов в форме нервных импульсов в мозг, который интерпретирует различия между колебаниями тканей тела и отолитов в качестве «звука». Величина и форма отолитов сильно различаются у разных видов, и это отражается на их способностях воспринимать колебательные возмущения в среде. Эти различия между видами обусловлены также свойствами другой структуры — именно, плавательного пузыря. Он наполнен газом, так что его плотность заметно ниже, чем у воды и тканей тела рыб. Поэтому пузырь может легко сжиматься в собственном ритме в ответ на колебания, поступающие извне. Если плавательный пузырь расположен в непосредственной близости от внутреннего уха, соединяется с ним системой контактирующих друг с другом косточек или даже примыкает к нему вплотную, то его колебания воздействую (через смещения отолитов) на чувствительные клетки внутри улитки. Те виды рыб, организм которых отвечает этим условиям, обладают достаточно высокими слуховыми способностями. Они могут воспринимать колебания с частотами 3 кГц и выше. В этом отношении они отличаются от видов, у которых плавательный пузырь отсутствует (акулы и скаты) или же помещается поодаль от внутреннего уха. У таких рыб способности слышать сильно ограничены, как и у придонных, обладающих маленьким плавательным пузырем. У них верхняя граница воспринимаемых частот не превышает 1 кГц. Другой инструмент восприятия колебаний у рыб — это так называемая боковая линия. Она представляет собой набор чувствительных структур, размещенных линейно по обеим сторонам головы и тела. Эти так называемые нейромасты локализованы либо на поверхности тела, либо в канале, который имеет выходы во внешнюю среду через поры, размещенные регулярно на одинаковых расстояниях друг от друга. Нейромаст построен из волосковых клеток того же характера, что находятся в улитке. Они отвечают на колебания частиц воды электрическими разрядами, поступающими в качестве сигналов в мозг рыбы. Боковая линия воспринимает только те колебания, источник которых находится не далее дистанции, равной длине тела рыбы, в лучшем случае — двух ее корпусов. Частоты таких колебаний не превышают 160—200 Гц. Именно работа боковой линии позволяет всем членам стаи рыб совершать во время ее движения на редкость синхронные маневры. Генерация звуков. Как писал Таволга, аппараты рыб, производящие звуки, настолько же разнообразны, насколько многообразны строение и образ жизни самих рыб (рис. 6.1). По мнению этого автора, едва ли удалось бы уложить различные типы звуковых механизмов рыб в рамки общей классификации эволюционных взаимосвязей между их семействами. Если оставить в стороне те волновые колебания воды, которые порождаются самими перемещениями рыбы[4], то все прочие механизмы можно разделить в грубой схеме на две категории. Это, во-первых, трение жестких структур друг о друга, за счет чего возникают механические звуки типа стридуляции. И, во-вторых, сокращения мышц, воздействующие на изменения конфигурации и объема плавательного пузыря. Что касается гидродинамических эффектов, вызванных плаванием рыб, то они относятся, скорее всего, к побочным эффектам их жизнедеятельности и, таким образом, могут, вероятно, иметь лишь косвенное отношение к коммуникации в строгом смысле этого слова. Их иногда называют звуками, издаваемыми «непреднамеренно». 2. Способы генерации звуковАкустическая коммуникация у рыб предполагает передачу акустических сигналов от одной особи вида к другой. Звуки как средство общения рыб чаще всего используются в контексте кормления, агрессии или ухаживания. Издаваемые звуки могут различаться в зависимости от вида и стимула. Рыбы могут издавать стридуляционные звуки, двигая компоненты скелетной системы, или могут издавать не стридуляторные звуки, манипулируя специализированными органами, такими как плавательный пузырь. Стридуляция. Наиболее распространенный способ генерации механических звуков — это трение друг о друга противолежащих участков так называемых глоточных зубов, которыми обладает большинство видов рыб. Известно, что скрежетание глоточными зубами наблюдается у представителей многих семейств костистых рыб. Почти любая рыба, которая питается крупной добычей, вероятно, издает звуки в процессе питания. Воздействие зубов на пищу даже средней твердости будет неизбежно порождать звуки. Челюстные зубы большинства видов представляют собой простые захватывающие устройствами, подобными клещам или ножницам. Но у некоторых видов они преобразовались в костные пластинки. У представителей отряда скалозубообразных (Tetraodontiformes), который включает в себя около 320 видов, обитающих в тропических и субтропических морских акваториях, зубы видоизменились либо в клюв, подобный клюву попугая, либо в дробящие пластинки. Эти создания могут издавать звуки путем трения фрагментов таких структур друг о друга. Важно заметить, что тем же способом пользуются морские караси из семейства Sparidae, совершенно неродственного скалозубообразным. Следует сказать, что скрежетание глоточными зубами наблюдается у этих и у других видов рыб не только во время поедания пищи. Механические звуки могут быть вызваны трением друг о друга более или менее видоизмененных лучей плавников. Лабиринтовая рыба Trichopsis vittatus (семейство Anabatidae) производит «бренчание» путем трения четвертого и пятого лучей грудных плавников о второй и третий лучи. Результатом может быть высокий по тону скрип, воспроизводимый, в частности, морским сомиком Galeichthys felis (семейство Ariidae). У спинорогов рода Batistes звуки воспроизводятся движением передних игл спинных плавников. У амфиприонов рода Amphiprion (семейство Pomacentridae), а также морских коньков Hippocampus и морских игл Syngnathus (семейство Cygnathidae) описаны звуки, порождаемые трением одной кости о другую. У хемулена Haemulon plumieri (семейство Pomadasyidae) генератором механических звуков оказываются жесткие структуры в основаниях жаберных дуг. Считают, что сигнал приобретает специфическое звучание за счет резонанса, создаваемого плавательным пузырем. Аналогичные явления имеют место у одного из видов спинорогов (род Badistes, семейство Balisti-dae) и неродственного ему бычка-подкаменщика (род Myoxocephalus, семейство Cottidae/ У них звуки порождаются щелканием сочленения между двумя костями грудного пояса. Этот глухой звук удара возникает при соприкосновении этих костей с передним концом плавательного пузыря, который, очевидно, выполняет роль резонатора. Генерация звуков плавательным пузырем. Понятно, что основная функция плавательного пузыря — это поддержание тела рыбы на плаву. Но он же, как мы видели, играет важную роль в восприятии волновых колебаний. Одновременно плавательный пузырь может служить не только в качестве резонатора, как у хемуленов и ряда других видов, но и аппаратом, генерирующим звуки. Это происходит за счет сокращения мышц, находящихся либо в стенках самого плавательного пузыря, либо соединенных одним концом с ним, а другим — прикрепленным к костям скелета. Сокращение мышц вызывает быстрое изменение объема плавательного пузыря, что и служит источником звука. Плавательный пузырь и его мускулатура, даже будучи извлечены из тела, могут служить эффективным устройством для изучения подводных звуков — стоит лишь заставить мышцы вибрировать под действием электрического стимулятора. К числу наиболее известных видов, которым свойственен такой тип генерации звуков, относятся опсанусы (род Opsanus, семейство Batrachoididae). У них плавательный пузырь выглядит как сердцевидное образование с закругленной вершиной, направленной назад. Две основные группы мышц, вызывающие изменения объема этого органа, крепятся к его боковой поверхности. Сходным образом устроен механизм порождения звуков у рыб других семейств, далеко отстоящего от опсанусов на эволюционной лестнице. Таковы, в частности, морские петухи из родов Trigla и Prionotus (семейство Triglidae). У рыб-барабанщиков из семейства горбылевых (Sciaenidae)[5] барабанные звуки производятся парой мышечных волокон, расположенных в боковых стенках тела. Они идут от массы спинных мышц, которые расположены почти вплотную к стенке плавательного пузыря. Результатом мышечных сокращений оказывается сжатие этого органа. Детали устройства этих мускулов и плавательного пузыря различны у разных видов, но их принципиальное строение сходно у всех тех многочисленных представителей названного семейства, которые способны издавать звуки. Важно заметить, что описанные особенности мускулатуры свойственны обычно только половозрелым самцам. Несколько по-иному, хотя и похожим образом, устроен аппарат генерации звуков у видов семейства ариевых (Ariidae) — морского сомика Galeichthys felis и косатки Bagre marinus. Парные короткие мышцы тянутся от верхних ушных костей в задней части черепа и проходят вдоль тонких, гибких костей — отростков первых четырех позвонков. Эти кости прилегают к верхней стенке плавательного пузыря. Сокращения названных мышц уравновешиваются эластичностью кромок этих костей. В результате быстрые серии мышечных сокращений вызывают вибрацию таких пружинистых образований и приводят к колебаниям газа, заполняющего плавательный пузырь. У южного атлантического побережья США ученые констатировали высокий уровень шумов, создаваемых большими косяками морских сомиков. Сходный механизм генерации звуков выработался у представителей двух других семейств, неродственных как ариевым, так и друг другу. Это групер Epinephelus striatus (семейство каменных окуней Serranidae) и рыба-белка Holocentrus rufus (семейство Holocentridae). У них две мышцы, участвующие в порождении звука, также берут начало от затылочной области черепа, но проходят не по отросткам позвонков, а вдоль нескольких первых пар брюшных ребер. Эти ребра оказываются уплощенными и словно бы врастают в верхнюю стенку плавательного пузыря. Предполагается, что такой механизм может быть обычным и у других видов двух названных семейств[6]. Еще один механизм генерации звуков, обнаруженный, в частности, у европейского угря Anguilla anguilla (семейство Anguillidae), состоит в том, что маленькие пузырьки газа с силой выдавливаются из плавательного пузыря в пищевод или в глотку. Возникающее при этом трение выливается в звук, усиливаемый резонаторными свойствами плавательного пузыря. Характеристики звуков. Временная организация и основные частотные параметры тех изученных звуков, которые генерируются с помощью плавательного пузыря, определяются ритмом сокращений связанных с ним мышц. Он, в свою очередь, зависит от частоты разрядов соответствующих моторных нейронов. Скорость следования разрядов положительно коррелирует с температурой воды. Для большинства изученных видов характерны импульсные звуки. Межвидовые различия затрагивают длительность пульсов (от 10 до 50 мс), а также их количество в одной посылке и ритм следования. При прослушивании через гидрофон эти акустические конструкции напоминают быстрые удары молотка о полый деревянный чурбан, барабанный бой, скрипение деревянных петель, кваканье либо визг пневматической дрели, сверлящей бетон. Все они имеют широкую полосу заполнения, лежащую в низкочастотной зоне спектра — обычно от 20 до 350 Гц, редко до 1200 Гц. Немногим изученным видам свойственны тональные звуки, напоминающие глухой свист. Например, у опсануса Opsanus notatus его частоты варьируют в пределах от 80 до 180 Гц. Понятно поэтому, что у большинства рыб наиболее чувствительная зона слуха лежит в области от 300 до 800 Гц, и мало существует видов (если они есть вообще), которые были бы способны обнаруживать звуки частотой свыше 3000 Гц. Основная частота звука зависит от размеров производящей его рыбы: в целом, чем крупнее размер особи, тем ниже частота и тем глуше звучит сигнал. Эта закономерность объясняется, как полагают, различиями в резонирующих свойствах плавательного пузыря у рыб разной величины[7]. Виды, которые генерируют звуки с помощью этого органа, издают звуки более громкие, чем те, для которых характерна стридуляция. Впрочем, как указывают эти авторы, звуки большинства видов, которые, в принципе, могут подавать сигналы с использованием плавательного пузыря, учеными не описаны. Это относится, в частности, ко всем видам глубоководных рыб. Эффективная зона распространения звуков. Эта сторона вопроса была изучена к концу прошлого века только лишь для мелководных рыб. При этом удалось установить, что низкочастотные звуки, то есть как раз те, которые наиболее свойственны рыбам и отвечают требованиям их слуховой системы, имеют в воде весьма узкую эффективную зону распространения. Например, при изучении степени деградации звуков, издаваемых зеленой рыбой-ласточкой Dascyllus albisella на глубине 7 м, оказалось, что большая часть параметров сигнала (длительность пульсов, основная частота и амплитудная модуляция пульсов внутри посылки) претерпевают существенные искажения уже на расстоянии 2 м от его отправителя. Лишь один показатель, именно период частоты следования пульсов, сохраняет относительное постоянство в радиусе до 11—12 м. Самцы африканской слабоэлектрической рыбы Pollimyrus isidori (семейство Mormyridae) издают два типа широкополосных звуков («ворчание» и «завывание»), адресуемых самкам. Несущая частота этих звуков — около 340 Hz, при верхней границе частотного диапазона около 3 kHz. Звуковое давление составляет 130 дб на расстоянии 10 см, но уже в полуметре от источника звука его акустические характеристики сильно искажаются. Это можно было бы рассматривать в качестве дефектности сигнала, если не принимать во внимание, что размер территории самца обычно не превышает 1 м в диаметре[8]. Эти и ряд других исследований заставляют сделать вывод, что коммуникация с использованием акустического канала у рыб возможна, в принципе, лишь на очень коротких дистанциях. Это связано еще и с тем, что на мелководье искажению сигналов способствует близость средового раздела вода-воздух, а на больших глубинах очень высок уровень фонового шума, производимого как беспозвоночными (см. главу 5), так и множеством видов рыб, живущих совместно. Иными словами, приходится прийти к заключению, что рыбы лишены возможности пользоваться чуть не главным преимуществом акустического канала связи. Я имею в виду возможность особи, живущей в местообитании с ограниченной видимостью, извещать о своем присутствии конспецификов, находящихся от нее на значительных расстояниях, и, соответственно, принимать звуковые дистантные сигналы, поступающие от прочих членов данной локальной популяции. Этот вывод подтверждается еще и тем обстоятельством, что рыбы способны определять местонахождение источников звука только в ближнем поле, но не способны делать это в масштабе значительных расстояний. О возможных коммуникативных функциях звуков рыб. Причины, побудившие биологов заняться вплотную изучением акустики рыб, оказались довольно неожиданными. Дело происходило в годы Второй мировой войны. В это время серьезной проблемой для военных моряков стали непонятные шумы, мешавшие, в частности, связи между судами. Вероятно, первыми об их возможном биологическом происхождении начали догадываться японцы, озабоченные защитой Токийского залива от управляемых акустических мин, которые могли быть использованы военными судами США. Те же непонятные явления беспокоили и американских моряков: в сообщениях подводных лодок, патрулирующих побережье США, часто содержались жалобы на необъяснимые «шипение» и «треск» в обширных мелководных районах, которые улавливались шумопеленгаторами, маскировали при этом все прочие шумы в воде и тем самым становились серьезной проблемой. В обеих воюющих странах по поручению командований их вооруженными силами начались всесторонние исследования источников подводных шумов, заметная доля которых оказалась звуками, производимыми рыбами. ВМС США установили гидрофоны вдоль всего юго-западного тихоокеанского побережья страны, а также вокруг Гавайских островов и других, находящихся в южной части Тихого океана. Интересно, что первые записи звуков, генерируемых морскими сомиками Galeichthys felis в естественных условиях обитания, были получены в 1943 г. Лабораторией оружия ВМС США у побережья штата Северная Каролина[9]. В последующие два-три десятилетия изучение голосов рыб проходило в основном по линии накопления сведений относительно их разнообразия и составления каталогов магнитофонных записей. Поскольку коллектирование вели, по сути дела «вслепую», с помощью гидрофонов, в этот период могли высказываться лишь догадки о возможной роли звуков в коммуникативном поведении рыб. Предположения о том, что акустический канал связи играет существенную роль в социальном и половом их поведении, основывались на данных по сезонной и суточной динамике интенсивностей акустической активности. Например, систематическая фиксация звуков нескольких (по крайней мере трех) видов рыб-барабанщиков, обитающих совместно у побережья США, показала, что в одном из районов эта активность быстро возрастала после середины мая, достигала пика в течение первых трех недель июня и затем постепенно снижалась. Подобная одновременная вокализация множества особей вошла с тех пор в научную литературу под наименованием «хоров». При прослушивании через гидрофон, звуки хора рыб-барабанщиков вызывают воспоминания об уровне шума в кузнечном цехе, оборудование которого находится на полном ходу, или шума авиационного мотора, работающего на расстоянии около 4.5 м от наблюдателя. Эти хоры, в данном случае, начинались ежедневно в примерно в 17 ч, когда шум становился различимым, достигали пика около 22 ч, а затем медленно затихали к 2 ч 30 мин. Все это давало основания предполагать, что хоры каким-то образом тесно вплетены во взаимодействия на почве репродукции. В отношении хоров морских сомиков вида Galeichthys felis, которые имеют место в августе у побережья Флориды, Таволга писал, что вызываемый ими шум напоминает «кипение гигантского кофейника», и что в нем, вероятно, участвуют сотни, а то и тысячи особей[10]. Все эти наблюдения наводили на мысль, что в «колониях» того или иного вида рыб эти звуки могут обозначать «страх, защиту от оппонента, выражение дружеских связей и т. д.». Осуществление реальных попыток понять, соответствуют ли эти предположения действительности, и если да, то в какой степени, стало возможным лишь после того, как поведением рыб вплотную занялись этологи, оснащенные современной аудио-видео аппаратурой. В качестве примера тех результатов, которые могут быть получены с применением этих методов, приведу результаты одного из недавних исследований на интересующую нас тему. Полевые наблюдения и эксперименты были проведены с обитателями коралловых рифов — рыбами-бабочками Chaetodon multicinctus (семейство Chaentodontidae). Этому виду свойственна моногамия, и оба члена супружеской пары охраняют от конспецификов границы своей кормовой территории. Т. Трикас с коллегами помещали в пределы территорий семнадцати пар банку с одной либо двумя живыми особями того же вида, что позволило описать в деталях поведение резидентной пары в момент охраны ею своего участка, а также реакции «пришельцев» на попытки хозяев изгнать их прочь (рис. 6.2). Примерно спустя 10 с после внедрения чужака хозяева приближались к банке и оставались на расстоянии менее метра от нее на протяжении 80 % времени всего эксперимента. В это время они пытались атаковать рыб, помещенных в резервуар. В статье детально описана моторика агонистического поведения: сближение с конкурентом и «прыжок» в воде с резким поворотом, после чего активная особь становится головой к оппоненту а затем поворачивается так, что вся его боковая поверхность, украшенная вертикальными полосами, оказывается обращенной в его сторону (рис. 6.25). В последовательность действий входят резкий удар хвостом (рис. 6.2Л) и растопыривание грудных плавников (рис. 6.25), а также приподнимание заднего верхнего плавника. Все эти двигательные реакции порождают гидродинамические колебания[11] воды с частотами менее 100 Гц. Помимо них авторы обнаружили 5 разных типов сигналов, которые они называют «акустическими» и «звуками», причем 4 из них производится синхронно с той или иной моторной акцией. Например, при ударе хвостом слышен короткий (154 мс) широкополосный щелчок со средним значением концентрации энергии 69 Гц (разброс 51—109 Гц). Короткий пульс со средней частотой 229 Hz слышен при оттопыривании грудных плавников. Во время прыжка рыба издает серию пульсов (4—9) с пиком частотного заполнения в зоне 466 Hz. Лишь один вариант звуков, который издавала только одна из рыб, помещенных в банку, если их там было две, не сопровождался какими-либо движениями из числа обозначенных выше. Это «хрюканье» с основной частотой 163 Гц.  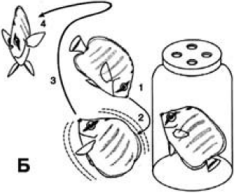   Рис. 2. Территориальное поведение рыб-бабочек Chaetodon multicinctus. Объяснения в тексте. Из: Tricas et al. 2006 Авторы работы констатируют использование рыбами-бабочками изученного вида «комлексных сигналов с оптическими, гидродинамическими и акустическими составляющими». Здесь перед нами вновь полимо-дальная конструкция, компоненты которой, транслируемые одновременно по нескольким каналам связи, разделимы лишь в грубой абстракции. По сути дела, каждая такая полимодальная акция представляет собой гештальт, который может быть воспринят адресатом сигнала как нечто единое и неразделимое целое[12]. Ф. Ладиш обобщил данные по использованию звуков в агонистических взаимодействиях (охрана территории, конфликты из-за доступа к корму и половому партнеру и т. д.) у 41 вида рыб из 29 семейств. Автор подчеркивает, что звуки почти всегда служат аккомпанементом к двигательным актам сигнального характера, что затрудняет выявление функционального значения акустического компонента. Так, например, у коралловой рыбы дасцилла трехполосого (Dascyllus aruanus, семейство Pomacentridae) самец приветствует самку, приплывшую к изготовленному им гнезду, характерным прыжком, Он взмывает вверх в толще воды, а затем устремляется вниз, одновременно издавая серию звуковых широкополосных импульсов (в среднем 5.4 в посылке) с несущей частотой около 370 Гц. По словам авторов статьи, то же самое поведение может служить «территориальным сигналом», поскольку в эксперимент, в котором самцу предъявляли особей того же пола, он воспроизводил сигналы того же характера. У цихловой рыбы Tramitichromis intermedius (семейство Cichlidae) участки самцов сгруппированы в плотные кластеры, и каждый строит в своих владениях гнездо, к которому пытается привлечь самок[13]. Этой цели служит акция, именуемая «дрожанием». Самец становится к самке боком, растопыривает непарные плавники, вибрирует задней частью туловища в горизонтальной плоскости с одновременными интенсивными боковыми движениями парных плавников. Акция оканчивается сильными боковыми ударами хвоста. Такое поведение приходится наблюдать все чаще с приближением сезона размножения. В 81 % случаев акция дрожания сопровождается генерацией звуков. Они воспроизводятся исключительно в ходе такого поведения. Однако тот факт, что лишь 19 % дрожаний проходят беззвучно, заставляет авторов предположить, что звук подается самцом преднамеренно, а не есть побочный механический эффект движений при дрожании. Впрочем, они сами указывают, что справедливость этой идеи следовало бы доказать дополнительно в последующих исследованиях. Репродуктивная биология другого вида цихловых рыб, Astato-tilapia burtoni аналогична тому, что описано для Т intermedius (скопления самцов, готовых к репродукции, вынашивание молоди во рту самок). Здесь звуки также ассоциируются с акциями дрожания. Звуки представляют собой пачки пульсов (2—19, в среднем 22 в посылке), с широким спектром частотного заполнения — от менее чем 50 Гц до 1.5 кГц, с несущей частотой около 22 Гц. Звучание сигнала тем глуше, чем крупнее размер самца. Авторы работы полагают, что эти звуки составляют важный компонент успеха самца у самок. В эксперименте самки значительно чаще нерестились с теми самцами, при контактах с которыми проигрывали записи звуков самцов, нежели с «молчаливыми» партнерами. Оказалось, что наравне с использованием модальностей оптической и акустической, в социальных отношениях между особями этого вида цихлид важную роль играет также и химическая. Самцы регулярно проделывают акты уринации при взаимодействии с самками (в особенности — с рецептивными) и с самцами-территориальными конкурентами. Частота этих актов возрастает у самца в 10 раз при виде самки, находящейся в состоянии готовности к икрометанию. Импульсы выбрасывания мочи не совпадают во времени с собственно половыми либо агонистическими взаимодействиями особей, а имеют место в коротких промежутках между такими контактами. Тем не менее, все то, что исследователи считают коммуникативными сигналами, имеет, как и во всех прочих ситуациях такого рода, сложнейшую полимодальную природу, никак не сводимую к действию сигналов какой-либо одной «преобладающей» модальности. 3. О чем говорят рыбы?О чем говорят рыбы? О еде и сексе! Они делают это на протяжении 155 миллионов лет и гораздо чаще, чем считается. К таким выводам пришли ученые из Корнеллского университета, пишет Science Daily. «Давно известно, что некоторые рыбы издают звуки. Но они всегда воспринимались, как редкость. Мы захотели узнать, действительно ли речь идет о единичных случаях, или у рыб существует широкий паттерн акустической коммуникации», — сказал ведущий автор исследования Аарон Райс. Он вместе с коллегами проанализировал данные о лучеперых рыбах. К этому классу относится больше 95% современных рыб. Оказалось, при помощи звуков общаются или имеют такую возможность 175 семейств — это две трети видов. Причем акустическая коммуникация настолько важна для рыб, что их звучание за миллионы лет эволюционировало 33 раза. В основном рыбы обсуждают друг с другом свое местоположение, защищают с помощью звука свою пищу и территорию или пытаются привлечь партнера. «Акустическая коммуникация рыб часто упускается из виду несмотря на то, что они составляют более половины всех ныне существующих видов позвоночных. Вероятно, дело в том, что рыбу нелегко услышать и даже увидеть, а наука о подводной акустической коммуникации в основном сосредоточена на китах и дельфинах. Но у рыб ведь тоже есть голоса!», — сказал соавтор исследования Эндрю Басс. В своей работе ученые использовали аудиозаписи со звуками рыб, научные статьи, их описывающие, и данные об анатомических особенностях. Теперь исследователи знают, как акустический функционал и коммуникативное поведение 35 тысяч известных видов рыб развивались на протяжении веков. ЗаключениеВ основном рыбы обсуждают друг с другом свое местоположение, защищают с помощью звука свою пищу и территорию или пытаются привлечь партнера. Анализ акустического поведения рыб позволяет увидеть в этих явлениях нечто общее с насекомыми. Там и тут, при колоссальной степени морфологической диверсификации видов, в каждом таксоне уровня рода или семейства независимо вырабатываются самые разные механизмы генерации звука. Они могут быть общими для большинства представителей данного таксона, либо варьировать в той или иной степени даже в его пределах. В то же время, похожие решения зачастую осуществляются у видов неродственных, далеко отстоящих друг от друга на эволюционной лестнице. Именно это мы видим у рыб. Так же как у насекомых (о чем было сказано в предыдущей главе), аппараты генерации звуков возникают, как полагают многие, на базе структур, которые первоначально служили целям оптимизации слуха, а отнюдь не потребностям внутривидовой коммуникации. Рыбы, в отличие от насекомых, находятся в двойственном положении в силу того, что обитают в среде с совершенно иными физическими свойствами. Дело в том, что самые разнообразные волновые возмущения в воде возникают вследствие передвижения в ней самой особи, как и любого «твердого тела». Эти возмущения позволительно назвать «звуками», лежащими в инфразвуковой зоне спектра. Такие возмущения можно подразделить на три основные типа: 1) более или менее ритмичные эффекты волнообразного движения; 2) турбулентность, порождаемую гидродинамическими шумами; 3) звуки, производимые изменениями напряжения внутренних органов при движении особи[14]. Наиболее интенсивные из этих эффектов наблюдаются тогда, когда животное быстро поворачивается или меняет скорость. Так, например, при повороте волновое возмущение происходит за счет того, что головная часть рыбы описывает круговую траекторию, используя хвост в качестве точки вращения. Очевидно, что даже прямолинейное передвижение рыбы вызывает горизонтальные перемещения среды, и эти перемещения будут ритмичными, поскольку ритмичны и движения тела рыбы. Поворот головы приводит к сильному перемещению среды. Возникают волны сжатия, обнаруживаемые большинством гидрофонов в качестве звуков. Иллюстрацией сказанному может служить картина агонистического поведения рыб-бабочек Chaetodon multicinctus (рис. 6.2). В этом случае сами по себе движения всего тела с резкими его поворотами, а так же вибрации тех или иных плавников генерируют акустические «сигналы» гидродинамического характера с собственными специфическими параметрами. Возникает вопрос, действительно ли звуковой аккомпанемент усиливает информативность этих гидродинамических сигналов, генераторы которых (сами движения рыбы и ее плавников) к тому же параллельно воспринимаются другим участником взаимодействия по оптическому каналу связи? Ответ будет положительным, если встать на широко распространенную точку зрения, согласно которой животные столь же хорошо распознают тонкие оттенки поведения своих социальных компаньонов, как это делает наблюдатель-исследователь. Я же придерживаюсь совершенно иных взглядов и полагаю, что звук в подобных случаях едва ли несет значимую функциональную нагрузку и может скорее рассматриваться в качестве некой избыточной компоненты (категория «непреднамеренных сигналов» или, точнее говоря, побочных механических эффектов). Сами конструктивные основы строения тела рыб не дают им возможности использовать те или иные структуры для генерации высокочастотных звуков. Это невыполнимо посредством стридуляции или экстренных изменений объема плавательного пузыря. Такого рода механизмы в состоянии порождать лишь низкочастотные звуки, которые в воде быстро дегенерируют и не могут, таким образом, служить инструментом дистантной акустической связи. Таким образом, на мой взгляд, акустическая компонента в тех сферах сигнального поведения рыб, которые регулируют половые и территориальные отношения, играет, в лучшем случае, совершенно подчиненную роль по сравнению с информацией, поступающей по химическому и оптическому каналам связи. Иное дело — гидродинамические эффекты, воспринимаемые органом боковой линии и обеспечивающие высокую согласованность траекторий направленного движения в мобильных стаях рыб. Этот тип анонимных связей между особями, обеспечивающий, к примеру, экстренное избегание хищников, можно было бы рассматривать в качестве слабой формы коммуникативного поведения, но вопрос этот следовало бы отнести к категории чисто терминологических. 1 Эндрюс Т. Язык животных. –М.: Крон-Пресс, 2000-607 с. |