бх. биохимия крови. 11. биохимия крови
 Скачать 261.07 Kb. Скачать 261.07 Kb.
|

|
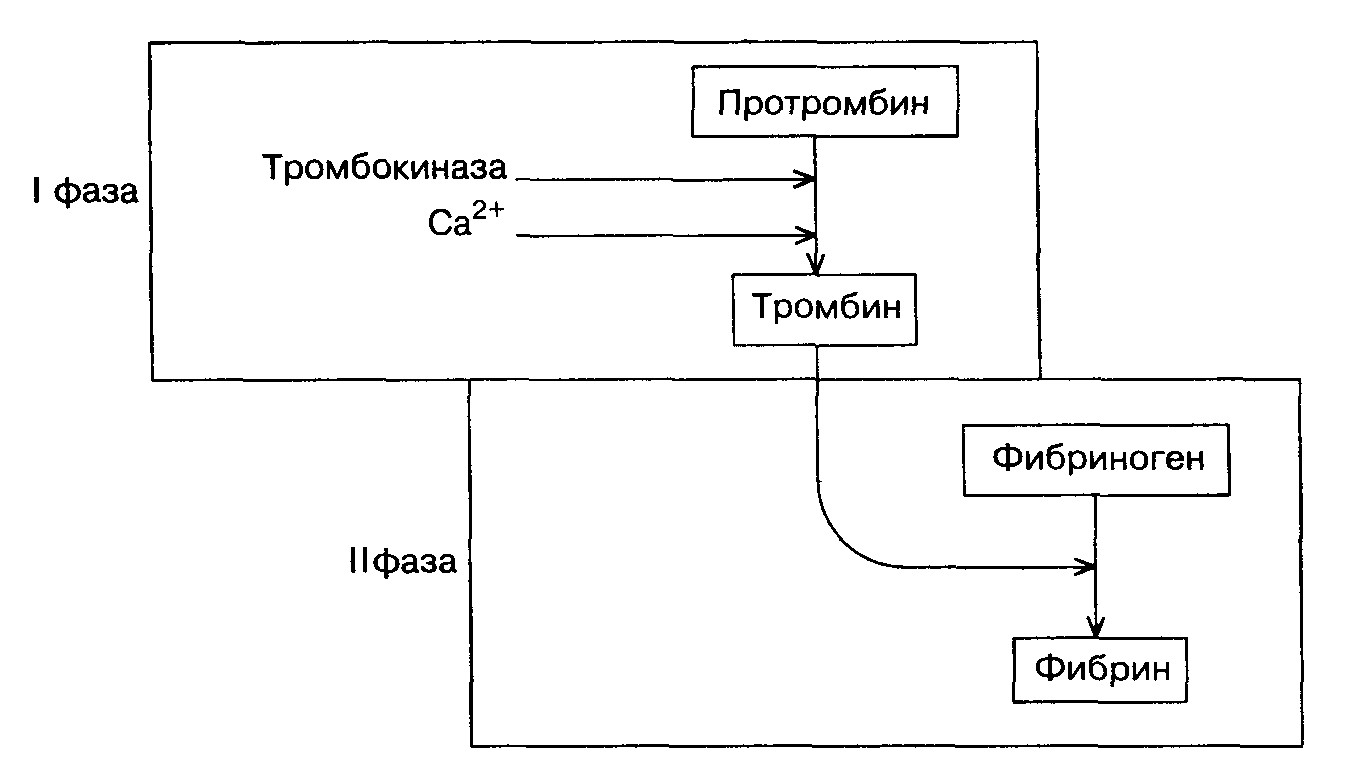
11. БИОХИМИЯ КРОВИ Главные функции крови связаны что тем, что это жидкая подвижная ткань, перемещающаяся по кровеносным сосудам. Кровь выполняет роль транспортного и коммуникативного средства интеграции обмена веществ разных органов. К основным функциям крови относятся: - дыхательная функция (перенос О2 из лёгких в ткани и СО2 из тканей в лёгкие); - трофическая функция (перенос продуктов пищеварения от кишечника к разным органам: глюкозы и кетоновых тел из печени в мышцы, жиров из печени в жировую ткань, молочной кислоты из мышц в печень, жирных кислот из жировой ткани в разные органы и т.д.); - выделительная функция (перенос мочевины из печени в почки, билирубина из разных тканей в печень и т.д.); - коммуникативная (регуляторная) функция (перенос гормонов и других регуляторных веществ к органам-мишеням). С движением крови связаны также и другие её функции: защитная, выполняемая антителами и фагоцитирующими лейкоцитами крови; участие в регуляции водно-солевого и кислотно-щелочного баланса; регуляция температуры тела путём теплообмена между тканями и движущейся кровью. Общее количество крови в организме взрослого человека составляет около 5 л (примерно 7% от массы тела). Она состоит из жидкой части и форменных элементов: эритроцитов, лейкоцитов и тромбоцитов, после осаждения которых, путём центрифугирования, над осадком остаётся светло-прозрачная жидкость – плазма крови. Белок плазмы фибриноген при стоянии превращается в нерастворимый фибрин, который образует сгусток. Из сгустка выжимается жидкость – сыворотка крови. Сыворотка отличается от плазмы тем, что в ней не содержится белок фибриноген. Состав плазмы является своего рода зеркалом метаболизма, поскольку изменения концентрации метаболитов в клетках, даже если они происходят в отдельных органах, отражаются на концентрации этих метаболитов в крови. Плазма крови представляет собой примерно 10% водный раствор органических и минеральных веществ. Концентрация белков составляет около 7%, минеральных солей около 1%, остальная часть приходится на различные небелковые органические соединения: мочевину, билирубин, индикан, креатин, креатинин, мочевую кислоту, глюкозу, фруктозу, молочную кислоту, пировиноградную кислоту и др Белки плазмы представлены пятью фракциями: альбуминами, α1-глобулинами, α2-глобулинами, β-глобулинами и -глобулинами. Каждая из этих фракций представляет собой смесь разных белков. Наиболее однородна альбуминовая фракция. Фракция -глобулинов содержит главным образом антитела (иммуноглобулины). Другие фракции гетерогенны. Большинство белков, содержащихся в плазме, синтезируются в печени. На долю альбуминов приходится более половины (55-60% или 40-50 г/л) белков плазмы крови. Альбумины выполняют функцию транспорта многих биологически активных веществ (гормонов, жирных кислот при мобилизации жиров из жировых депо). Они способны связываться с холестерином, желчными пигментами. Значительная часть кальция в сыворотке крови также связана с альбуминами. Альбумины играют важную роль в поддержании онкотического давления крови. Концентрация альбуминов в сыворотке ниже 30 г/л вызывает значительные изменения онкотического давления крови, что приводит к возникновению отёков. Концентрация альбумина в крови может снижаться вследствие его выделения с мочой при заболеваниях почек (альбуминурия). Печень человека синтезирует и выделяет в кровь до 15 г альбумина в сутки. При некоторых болезнях печени (циррозе) синтез альбумина нарушается. Для этих состояний тоже характерны отёки. Глобулины представлены 2 фракциями – эуглобулинами и псевдоглобулинами. Фракция эуглобулинов в основном состоит из -глобулинов, а фракция псевдоглобулинов из α- ,β- и -глобулинов. Большая часть антител находится во фракции -глобулинов. Иммуноглобулины, или антитела синтезируются β-лимфоцитами или образующимися из них плазматическими клетками. При ряде патологических состояний может наблюдаться гиперпротеинемия, обусловленная увеличением уровня -глобулинов: например, в результате инфекционного процесса. При многих заболеваниях изменяется процентное соотношение отдельных белковых фракций плазмы крови, хотя общее содержание белка в сыворотке крови остаётся в пределах нормы. Такое состояние носит название диспротеинемия. 11.1. ДЫХАТЕЛЬНАЯ ФУНКЦИЯ КРОВИ. БУФЕРНАЯ СИСТЕМА КРОВИ Кровь играет определяющую роль в поддержании кислотно-щелочного равновесия, изменение которого может привести к развитию патологических состояний или гибели организма. Поэтому в организме существуют специальные системы, которые препятствуют изменению рН крови и других биологических жидкостей при образовании кислых и щелочных продуктов или при большом поступлении воды. Такую роль выполняют отдельные физиологические системы (дыхательная, выделительная), а также буферные системы. Последние очень быстро (в течение нескольких секунд) реагируют на изменение концентрации Н+ и ОН- в водных средах и являются срочными регуляторами кислотно-основного состояния в тканях организма. Буферные системы – это смесь слабой кислоты и её растворимой соли, двух солей или белков, которые способны препятствовать изменению рН водных сред. Действие буферных систем направлено на связывание избытка Н+ или ОН- в среде и поддержание постоянства рН среды. При действии буферной системы образуются слабодиссоциируемые вещества или вода. К основным буферным системам крови относятся бикарбонатная, белковая (гемоглобиновая) и фосфатная. Имеются также ацетатная и аммонийная буферные системы. Бикарбонатная буферная система - мощная и самая управляемая система крови и внеклеточной жидкости. На её долю приходится около 10% всей буферной ёмкости крови. Бикарбонатная система представляет собой сопряжённую кислотно-основную пару, состоящую из молекулы угольной кислоты Н2СО3, выполняющую роль донора протона, и бикарбонат-иона НСО3-, выполняющего роль акцептора протона: СО2 + Н2О ↔ Н2СО3 ↔ Н+ + НСО3- Истинная концентрация недиссоциированных молекул Н2СО3 в крови незначительна и находится в прямой зависимости от концентрации растворённого СО2. При нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО3- в плазме крови превышает концентрацию СО2 примерно в 20 раз. Бикарбонатная буферная система функционирует как эффективный регулятор в области рН = 7,4. Механизм действия этой системы заключается в том, что при выделении в кровь относительно больших количеств кислых продуктов протоны Н+ взаимодействуют с ионами бикарбоната НСО3- , что приводит к образованию слабодиссоциируемой Н2СО3. Последующее снижение концентрации Н2СО3 достигается в результате ускоренного выделения СО2 через лёгкие в результате их гипервентиляции. Если в крови увеличивается количество оснований, то они, взаимодействуя со слабой угольной кислотой, образуют ионы бикарбоната и воду. При этом не происходит сколько-нибудь заметных сдвигов в величине рН. Кроме того, для сохранения нормального соотношения между компонентами буферной системы в этом случае подключаются физиологические механизмы регуляции кислотно-основного равновесия: происходит задержка в плазме крови некоторого количества СО2 в результате гиповентиляции лёгких. Бикарбонатная система тесно связана с гемоглобиновой системой. Фосфатная буферная система представляет собой сопряжённую кислотно-основную пару, состоящую из иона Н2РО4- (донор протонов, выполняет роль кислоты) и иона НРО42- (акцептор протонов, выполняет роль соли). Фосфатная буферная система составляет лишь 1% от буерной ёмкости крови. В других тканях эта система является одной из основных. Фосфатная буферная система способна оказывать влияние при изменениях рН в интервале от 6,1 до 7,7 и может обеспечивать определённую ёмкость внутриклеточной жидкости, величина рН которой в пределах 6,9-7,4. В крови максимальная ёмкость фосфатного буфера проявляется вблизи значения 7,2. Органические фосфаты также обладают буферными свойствами, но мощность их слабее, чем неорганического фосфатного буфера. Белковая буферная система имеет меньшее значение для поддержания кислотно-основного равновесия в плазме крови, чем другие буферные системы. Белки образуют буферную систему благодаря наличию кислотно-основных групп в молекуле белков: белок-Н+ (кислота, донор протонов) и белок (сопряжённое основание, акцептор протонов). Белковая буферная система плазмы крови эффективна в области значений рН 7,2-7,4. Гемоглобиновая буферная система – самая мощная буферная система крови, на её долю приходится 75% от всей буферной. Участие гемоглобина в регуляции рН крови связано с его ролью в транспорте кислорода и углекислого газа. При насыщении кислородом гемоглобин становится более сильной кислотой (ННbО2). Гемоглобин, отдавая кислород, превращается в очень слабую органическую кислоту (ННb). Гемоглобиновая буферная система состоит из неионизированного гемоглобина ННb (слабая органическая кислота, донор протонов) и калиевой соли гемоглобина КНb (сопряжённое основание, акцептор протонов). Точно так же может быть рассмотрена оксигемоглобиновая буферная система. Система гемоглобина и система оксигемоглобина являются взаимопревращающимися системами и существуют как единое целое. Буферные свойства гемоглобина обусловлены возможностью взаимодействия кисло реагирующих соединений с калиевой солью гемоглобина: КНb + Н2СО3 => КНСО3 + ННb. Это обеспечивает поддержание рН крови в пределах физиологически допустимых величин, несмотря на поступление в венозную кровь большого количества СО2 и других продуктов обмена кислотного характера. Гемоглобин (ННb), попадая в капилляры лёгких, превращается в оксигемоглобин (ННbО2), что приводит к некоторому подкислению крови, вытеснению части Н2СО3 из бикарбонатов и понижению щелочного резерва крови. Дыхательная функция крови. Важной функцией крови является её способность переносить кислород к тканям и СО2 от тканей к лёгким. Веществом, осуществляющим эту функцию, является гемоглобин. Гемоглобин способен поглощать О2 при сравнительно высоком содержании его в атмосферном воздухе и легко отдавать при понижении парциального давления О2: Нb + О2 ↔ НbО. Поэтому в легочных капиллярах происходит насыщение крови О2, в то время как в тканевых капиллярах, где парциальное давление его резко снижается, наблюдается обратный процесс – отдача кровью кислорода тканям. Образующийся в тканях при окислительных процессах СО2 подлежит выведению из организма. Обеспечение такого газообмена осуществляется несколькими системами организма. Наибольшее значение имеют внешнее, или легочное, дыхание, обеспечивающее направленную диффузию газов через альвеолокапиллярные перегородки в легких и обмен газов между наружным воздухом и кровью; дыхательная функция крови, зависимая от способности плазмы растворять и способности гемоглобина обратимо связывать кислород и углекислый газ; транспортная функция сердечно-сосудистой системы (кровотока), обеспечивающая перенос газов крови от легких к тканям и обратно; функция ферментных систем, обеспечивающая обмен газов между кровью и клетками тканей, т.е. тканевое дыхание. Диффузия газов крови осуществляется через мембрану клеток по концентрационному градиенту. За счет этого процесса в альвеолах легких в конце вдоха происходит выравнивание парциальных давлений различных газов в альвеолярном воздухе и крови. Обмен с атмосферным воздухом в процессе последующих выдоха и вдоха вновь приводит к различиям концентрации газов в альвеолярном воздухе и в крови, в связи с чем происходит диффузия кислорода в кровь, а углекислого газа из крови. Большая часть О2 и СО2 переносится в форме связи их с гемоглобином в виде молекул HbO2 и HbCO2. Максимальное количество кислорода, связываемое кровью при полном насыщении гемоглобина кислородом, называется кислородной емкостью крови. В норме ее величина колеблется в пределах 16,0–24,0 об.% и зависит от содержания в крови гемоглобина, 1 г которого может связать 1,34 мл кислорода (число Хюфнера). Связывание кислорода гемоглобином является обратимым процессом, зависимым от напряжения кислорода в крови, а также от других факторов, в частности от рН крови. СО2, образующийся в тканях, переходит в кровь кровеносных капилляров, затем диффундирует внутрь эритроцита, где под влиянием карбоангидразы превращается в угольную кислоту, которая диссоциирует на Н+ и НСО3-. НСО3- частично диффундируют в плазму крови, образуя бикарбонат натрия. Он при поступлении крови в легкие (как и ионы НСО3-, содержащиеся в эритроцитах) образует СО2, который диффундирует в альвеолы. Около 80% всего количества СО2 переносится от тканей к легким в виде бикарбонатов, 10% – в виде свободно растворенной углекислоты и 10% – в виде карбоксигемоглобина. Карбоксигемоглобин диссоциирует в легочных капиллярах на гемоглобин и свободный СО2, который удаляется с выдыхаемым воздухом. Освобождению СО2 из связи с гемоглобином способствует превращение последнего в оксигемоглобин, который, обладая выраженными кислотными свойствами, способен переводить бикарбонаты в угольную кислоту, диссоциирующую с образованием молекул воды и СО2. При недостаточном насыщении крови кислородом развивается гипоксемия, которая сопровождается развитием гипоксии, т.е. недостаточным снабжением тканей кислородом. Тяжелые формы гипоксемии могут вызвать полное прекращение доставки кислорода тканям, тогда развивается аноксия, в этих случаях наступает потеря сознания, которая может закончиться смертью. Патология газообмена, связанная с нарушением транспорта газов между легкими и клетками организма, наблюдается при уменьшении газовой емкости крови вследствие недостатка или качественных изменений гемоглобина, проявляется в виде анемических гипоксий. При анемиях кислородная емкость крови уменьшается пропорционально снижению концентрации гемоглобина. Снижение концентрации гемоглобина при анемиях ограничивает и транспорт углекислоты от тканей к легким в форме карбоксигемоглобина. Нарушение транспорта кислорода кровью возникает также при патологии гемоглобина, например при серповидно-клеточной анемии, при инактивации части молекул гемоглобина за счет превращения его в метгемоглобин, например, при отравлении нитратами (метгемоглобинемия), или в карбоксигемоглобин (отравление СО). Нарушения газообмена вследствие уменьшения объемной скорости кровотоки в капиллярах возникают при сердечной недостаточности, сосудистой недостаточности (в т.ч. при коллапсе, шоке), локальные нарушения – при ангиоспазме и др. В условиях застоя крови концентрация восстановленного гемоглобина возрастает. При сердечной недостаточности этот феномен особенно выражен в капиллярах отдаленных от сердца участков тела, где кровоток наиболее замедлен, что клинически проявляется акроцианозом. Первичное нарушение газообмена на уровне клеток наблюдается главным образом при воздействии ядов, блокирующих дыхательные ферменты. В результате клетки утрачивают способность утилизировать кислород, и развивается резкая тканевая гипоксия, приводящая к структурной дезорганизации субклеточных и клеточных элементов, вплоть до некроза. Нарушению клеточного дыхания может способствовать витаминная недостаточность, например дефицит витаминов В2, РР, являющихся коферментами дыхательных ферментов. 11.2. СИСТЕМА СВЕРТЫВАНИЯ КРОВИ. ИЗМЕНЕНИЯ ПРИ ПАТОЛОГИИ При случайных повреждениях мелких кровеносных сосудов возникающее кровотечение через некоторое время прекращается. Это связано с образованием в месте повреждения сосуда тромба или сгустка. Данный процесс называется свёртыванием крови. В настоящее время существует классическая ферментативная теория свертывания крови – теория Шмидта – Моравица. Положения этой теории представлены на схеме (рис. 11):
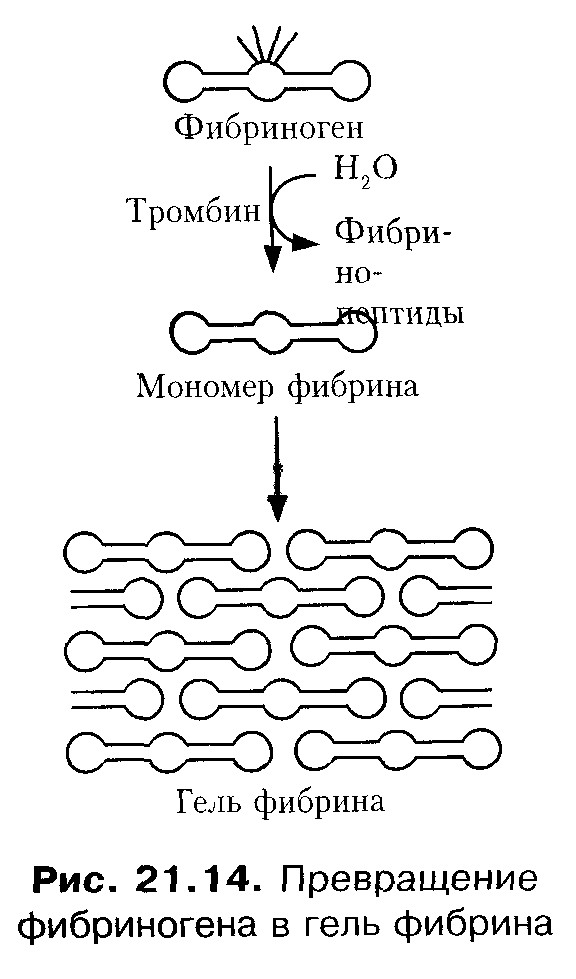
Повреждение кровеносного сосуда вызывает каскад молекулярных процессов, в результате образуется сгусток крови - тромб, прекращающий вытекание крови. В месте повреждения к открывшемуся межклеточному матриксу прикрепляются тромбоциты; возникает тромбоцитарная пробка. Одновременно включается система реакций, ведущих к превращению растворимого белка плазмы фибриногена в нерастворимый фибрин, который откладывается в тромбоцитарной пробке и на её поверхности, образуется тромб. Процесс свёртывания крови протекает в две фазы. В первой фазе протромбин переходит в активный фермент тромбин под влиянием тромбокиназы, содержащейся в тромбоцитах и освобождающейся из них при разрушении кровяных пластинок, и ионов кальция. Во второй фазе под влиянием образовавшегося тромбина фибриноген превращается в фибрин. Весь процесс свёртывания крови представлен следующими фазами гемостаза: а) сокращение поврежденного сосуда; б) образование в месте повреждения рыхлой тромбоцитарной пробки, или белого тромба. Коллаген сосуда служит связующим центром для тромбоцитов. При агрегации тромбоцитов освобождаются вазоактивные амины, которые стимулируют сужение сосудов; в) формирование красного тромба (кровяной сгусток); г) частичное или полное растворение сгустка. Белый тромб образуется из тромбоцитов и фибрина; в нем относительно мало эритроцитов (в условиях высокой скорости кровотока). Красный тромб состоит из эритроцитов и фибрина (в областях замедленного кровотока). В процессе свертывания крови участвуют факторы свертывания крови. Факторы свертывания, связанные с тромбоцитами, принято обозначать арабскими цифрами (1, 2, 3 и т.д.), а факторы свертывания, находящиеся в плазме крови, обозначают римскими цифрами. Фактор I (фибриноген) - гликопротеин. Синтезируется в печени. Фактор II (протромбин) - гликопротеин. Синтезируется в печени при участии витамин К. Способен связывать ионы кальция. При гидролитическом расщеплении протромбина образуется активный фермент свертывания крови. Фактор III (тканевый фактор, или тканевый тромбопластин) образуется при повреждении тканей. Липопротеин. Фактор IV (ионы Са2+). Необходимы для образования активного фактора X и активного тромбопластина тканей, активации проконвертина, образования тромбина, лабилизации мембран тромбоцитов. Фактор V (проакцелерин) - глобулин. Предшественник акцелерина, синтезируется в печени. Фактор VII (антифибринолизин, проконвертин)- предшественник конвертина. Синтезируется в печени при участии витамина К. Фактор VIII (антигемофильный глобулин А) необходим для формирования активного фактора X. Врожденный недостаток фактора VIII - причина гемофилии А. Фактор IX (антигемофильный глобулин В, Кристмас-фактор) принимает участие в образовании активного фактора X. При недостаточностьи фактора IX развивается гемофилия В. Фактор X (фактор Стюарта-Прауэра) - глобулин. Фактор X участвует в образовании тромбина из протромбина. Синтезируется клетками печени при участии витамина К. Фактор XI (фактор Розенталя) - антигемофильный фактор белковой природы. Недостаточность наблюдается при гемофилии С. Фактор XII (фактор Хагемана) участвует в пусковом механизме свертывания крови, стимулирует фибринолитическую активность, другие защитные реакции организма. Фактор XIII (фибринстабилизирующий фактор) - участвует в образовании межмолекулярных связей в фибрин-полимере. Факторы тромбоцитов. В настоящее время известно около 10 отдельных факторов тромбоцитов. Например: Фактор 1 - адсорбированный на поверхности тромбоцитов проакцелерин. Фактор 4 - антигепариновый фактор. В нормальных условиях тромбина в крови нет, он образуется из белка плазмы протромбина под действием протеолитического фермента фактора Ха (индекс а - активная форма), который образуется при кровопотере из фактора X. Фактор Ха превращает протромбин в тромбин только в присутствии ионов Са2 + и других факторов свертывания. Фактор III, переходящий в плазму крови при повреждении тканей, и фактор 3 тромбоцитов создают предпосылки для образования затравочного количества тромбина из протромбина. Он катализирует превращение проакцелерина и проконвертина в акцелерин (фактор Va) и в конвертин (фактор VIIa). При взаимодействии перечисленных факторов, а также ионов Са2+ происходит образование фактора Ха. Затем происходит образование тромбина из протромбина. Под влиянием тромбина от фибриногена отщепляются 2 пептида А и 2 пептида В. Фибриноген превращается в хорошо растворимый фибрин-мономер, который быстро полимеризуется в нерастворимый фибрин-полимер при участии фибринстабилизирующего фактора- фактора XIII (фермент трансглутаминаза) в присутствии ионов Са2+ (рис. 12).
Фибриновый тромб прикрепляется к матриксу в области повреждения сосуда при участии белка фибронектина. Вслед за образованием нитей фибрина происходит их сокращение, для чего необходима энергия АТФ и фактор 8 тромбоцитов (тромбостенин). У людей с наследственными дефектами трансглутаминазы кровь свертывается так же, как у здоровых, однако тромб получается хрупкий, поэтому легко возникают вторичные кровотечения. Кровотечение из капилляров и мелких сосудов останавливается уже при образовании тромбоцитной пробки. Для остановки кровотечения из более крупных сосудов необходимо быстрое образование прочного тромба, чтобы свести к минимуму потерю крови. Это достигается каскадом ферментных реакций с механизмами усиления на многих ступенях. Различают три механизма активации ферментов каскада: 1. Частичный протеолиз. 2. Взаимодействие с белками-активаторами. 3. Взаимодействие с клеточными мембранами. Ферменты прокоагулянтного пути содержат γ-карбоксиглутаминовую кислоту. Радикалы карбоксиглутаминовой кислоты образуют центры связывания ионов Са2+. В отсутствие ионов Са2+ кровь не свертывается. Внешний и внутренний пути свёртывания крови. Во внешнем пути свертывания крови участвуют тромбопластин (тканевой фактор, фактор III), проконвертин (фактор VII), фактор Стюарта (фактор X), проакцелерин (фактор V), а также Са2+ и фосфолипиды мембранных поверхностей, на которых образуется тромб. Гомогенаты многих тканей ускоряют свёртывание крови: это действие называют тромбопластиновой активностью. Вероятно, она связана с наличием в тканях какого-то специального белка. Факторы VII и X - проферменты. Они активируются путём частичного протеолиза, превращаясь в протеолитические ферменты - факторы VIIа и Xа соответственно. Фактор V – это белок, который при действии тромбина превращается в фактор V', который не является ферментом, но активирует фермент Xа по аллостерическому механизму; активация усиливается в присутствии фосфолипидов и Са2+. В плазме крови постоянно содержатся следовые количества фактора VIIа. При повреждении тканей и стенок сосуда освобождается фактор III – мощный активатор фактора VIIа; активность последнего увеличивается более чем в 15000 раз. Фактор VIIа отщепляет часть пептидной цепи фактора X, превращая его в фермент - фактор Xа. Сходным образом Xа активирует протромбин; образовавшийся тромбин катализирует превращение фибриногена в фибрин, а также превращение предшественника трансглутаминазы в активный фермент (фактор XIIIа). Этот каскад реакций имеет положительные обратные связи, усиливающие конечный результат. Фактор Xа и тромбин катализируют превращение неактивного фактора VII в фермент VIIа; тромбин превращает фактор V в фактор V', который вместе с фосфолипидами и Са2+ в 104–105 раз повышает активность фактора Xа. Благодаря положительным обратным связям скорость образования самого тромбина и, следовательно, превращения фибриногена в фибрин нарастают лавинообразно, и в течение 10-12 с кровь свёртывается. Свёртывание крови по внутреннему механизму происходит значительно медленнее и требует 10-15 мин. Этот механизм называют внутренним, потому что для него не требуется тромбопластин (тканевой фактор) и все необходимые факторы содержатся в крови. Внутренний механизм свёртывания также представляет собой каскад последовательных активаций проферментов. Начиная со стадии превращения фактора X в Xа, внешний и внутренний пути одинаковы. Как и внешний путь, внутренний путь свёртывания имеет положительные обратные связи: тромбин катализирует превращение предшественников V и VIII в активаторы V' и VIII', которые в конечном итоге увеличивают скорость образования самого тромбина. Внешний и внутренний механизмы свёртывания крови взаимодействуют между собой. Фактор VII, специфичный для внешнего пути свёртывания, может быть активирован фактором XIIа, который участвует во внутреннем пути свёртывания. Это превращает оба пути в единую систему свёртывания крови. Гемофилии. Наследственные дефекты белков, участвующих в свёртывании крови, проявляются повышением кровоточивости. Наиболее часто встречается болезнь, вызванная отсутствием фактора VIII – гемофилия А. Ген фактора VIII локализован в X- хромосоме; повреждение этого гена проявляется как рецессивный признак, поэтому у женщин гемофилии А не бывает. У мужчин, имеющих одну X-хромосому, наследование дефектного гена приводит к гемофилии. Признаки болезни обычно обнаруживаются в раннем детстве: при малейшем порезе, а то и спонтанно возникают кровотечения; характерны внутрисуставные кровоизлияния. Частая потеря крови приводит к развитию железодефицитной анемии. Для остановки кровотечения при гемофилии вводят свежую донорскую кровь, содержащую фактор VIII, или препараты фактора VIII. Гемофилия В. Гемофилия В обусловлена мутациями гена фактора IX, который, как и ген фактора VIII, локализован в половой хромосоме; мутации рецессивны, следовательно, гемофилия В бывает только у мужчин. Гемофилия В встречается примерно в 5 раз реже, чем гемофилия А. Лечат гемофилию В введением препаратов фактора IX. При повышенной свертываемости крови могут образоваться внутрисосудистые тромбы, закупоривающие неповрежденные сосуды (тромботические состояния, тромбофилии). Фибринолиз. Тромб в течение нескольких дней после образования рассасывается. Главная роль в его растворении принадлежит протеолитическому ферменту плазмину. Плазмин гидролизирует в фибрине пептидные связи, образованные остатками аргинина и триптофана, причём образуются растворимые пептиды. В циркулирующей крови находится предшественник плазмина – плазминоген. Он активируется ферментом урокиназой, который содержится во многих тканях. Пламиноген может активироваться калликреином, также имеющимся в тромбе. Плазмин может активироваться и в циркулирующей крови без повреждения сосудов. Там плазмин быстро инактивируется белковым ингибитором α2- антиплазмином, в то время как внутри тромба он защищён от действия ингибитора. Урокиназа – эффективное средство для растворения тромбов или предупреждения их образования при тромбофлебитах, тромбоэмболии легочных сосудов, инфаркте миокарда, хирургических вмешательствах. Противосвёртывающая система. При развитии системы свёртывания крови в ходе эволюции решались две противоположные задачи: предотвращать вытекание крови при повреждении сосудов и сохранять кровь в жидком состоянии в неповреждённых сосудах. Вторая задача решается противосвёртывающей системой, которая представлена набором белков плазмы, ингибирующих протеолитические ферменты. Белок плазмы антитромбин III ингибирует все протеиназы, участвующие в свёртывании крови, кроме фактора VIIа. Он не действует на факторы, находящиеся в составе комплексов с фосфолипидами, а только на те, которые находятся в плазме в растворённом состоянии. Следовательно, он нужен не для регуляции образования тромба, а для устранения ферментов, попадающих в кровоток из места образования тромба, тем самым он предотвращает распространение свёртывания крови на поврежденные участки кровеносного русла. В качестве препарата, предотвращающего свёртывание крови, применяется гепарин. Гепарин усиливает ингибирующее действие антитромбина III: присоединение гепарина индуцирует конформационные изменения, которые повышают сродство ингибитора к тромбину и другим факторам. После соединения этого комплекса с тромбином гепарин освобождается и может присоединяться к другим молекулам антитромбина III. Таким образом, каждая молекула гепарина может активировать большое количество молекул антитромбина III; в этом отношении действие гепарина сходно с действием катализаторов. Гепарин применяют как антикоагулянт при лечении тромботических состояний. Известен генетический дефект, при котором концентрация антитромбина III в крови вдвое меньше, чем в норме; у таких людей часто наблюдаются тромбозы. Антитромбин III – главный компонент противосвёртывающей системы. В плазме крови есть и другие белки – ингибиторы протеиназ, которые также могут уменьшать вероятность внутрисосудистого свёртывания крови. Таким белком является α2- макроглобулин, который ингибирует многие протеиназы, и не только те, которые участвуют в свёртывании крови. α2-Макроглобулин содержит участки пептидной цепи, которые являются субстратами многих протеиназ; протеиназы присоединяются к этим участкам, гидролизируют в них некоторые пептидные связи, в результате чего изменяется конформация α2-макроглобулина, и он захватывает фермент, подобно капкану. Фермент при этом не повреждается: в комплексе с ингибитором он способен гидролизировать низкомолекулярные пептиды, но для крупных молекул активный центр фермента не доступен. Комплекс α2-макроглобулина с ферментом быстро удаляется из крови: время его полужизни в крови около 10 мин. При массивном поступлении в кровоток активированных факторов свёртывания крови мощность противосвёртывающей системы может оказаться недостаточной, и появляется опасность тромбозов. Витамин К. В пептидных цепях факторов II, VII, IX, и X содержится необычная аминокислота - γ-карбоксиглутаминовая. Эта аминокислота образуется из глутаминовой кислоты в результате посттрансляционной модификации указанных белков: 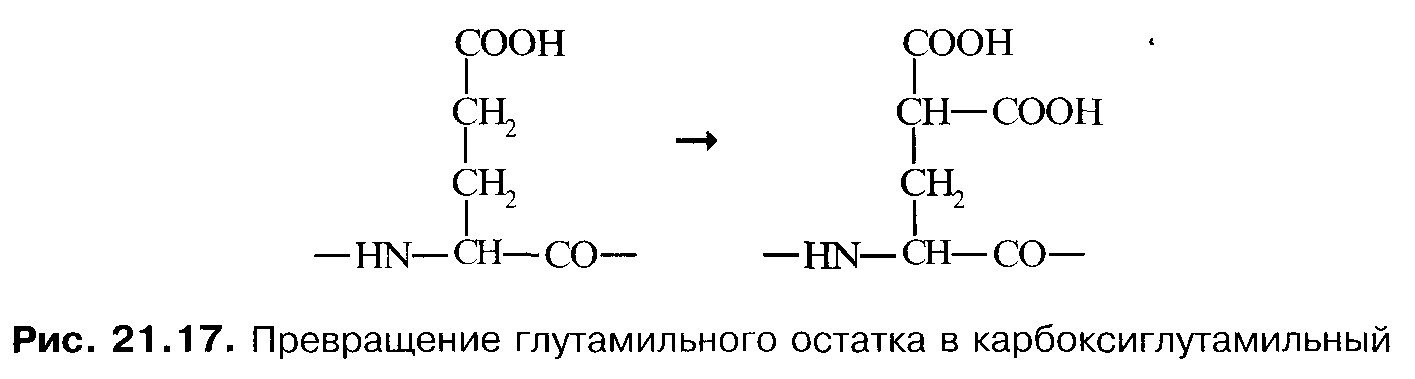 Реакции, в которых участвуют факторы II, VII, IX, и X, активируются ионами Са2+ и фосфолипидами: радикалы γ-карбоксиглутаминовой кислоты образуют центры связывания Са2+ на этих белках. Перечисленные факторы, а также факторы V' и VIII' прикрепляютя к бислойным фосфолипидным мембранам и друг к другу при участии ионов Са2+, и в таких комплексах происходит активация факторов II, VII, IX, и X. Ион Са2+ активирует также и некоторые другие реакции свёртывания: декальцинированная кровь не свёртывается. Превращение глутамильного остатка в остаток γ-карбоксиглутаминовой кислоты катализируется ферментом, коферментом которого служит витамин К. Недостаточность витамина К проявляется повышенной кровоточивостью, подкожными и внутренними кровоизлияниями. В отсутствие витамина К образуются факторы II, VII, IX, и X, не содержащие γ-карбоксиглутаминовых остатков. Такие проферменты не могут превращаться в активные ферменты. |