Отражатель игк 30б. Лекция 9. 2. Краткие сведения о геохронологии Возникновение жизни. Основные этапы биогенеза
 Скачать 0.84 Mb. Скачать 0.84 Mb.
|

|
Происхождение и развитие жизни на Земле 1. Жизнь как особая форма движения материи. Свойства и уровни организации живого. Гипотезы происхождения жизни 2. Краткие сведения о геохронологии 3. Возникновение жизни. Основные этапы биогенеза 4. Эволюция одноклеточных организмов 5. Возникновение и развитие многоклеточной организации 6. Эволюция животного мира 7. Эволюция растительного мира Жизнь как особая форма движения материи. Свойства и уровни организации живого. Гипотезы происхождения жизни. Жизнь – одно из сложнейших явлений природы. В XVII-XVIII вв. широкое распространение получил витализм (от лат. vitalis - жизненный), основоположником которого считают древнегреческого философа Аристотеля. По Аристотелю «жизнь есть питание, рост и одряхление, причиной которых является энтелехия – принцип, имеющий цель в самом себе». Сторонники витализма предполагали, что организмам присуща особая «жизненная сила» или «души», которая управляет всеми жизненными процессами. Как только она покидает тело, организм начинает разрушаться. Жизненная сила, как особая нематериальная субстанция, считалась независимой от материи. Поэтому последовательные сторонники виталистической концепции считали ее проявлением божественного начала. Виталисты считали, что живые организмы состоят из органических веществ, которые невозможно получить искусственным путем, что к живым организмам неприменим закон сохранения энергии. Однако эти утверждения были опровергнуты немецким химиком Ф. Велером, который в 1828 г. впервые синтезировал в лабораторных условиях органическое вещество (мочевину). В настоящее время искусственным путем получено свыше 100000 органических веществ. В XVIII в. был распространен механистический взгляд на природу, в соответствии с которым живые организмы рассматривались как особые механизмы, отличающиеся от созданных человеком только сложностью строения. Такая концепция оказалось бессильной решить проблему сущности жизни, так как создатели ее не видели специфики живых тел природы. В дальнейшем попытки определить сущность жизни также предпринимались неоднократно. Павлов определяет жизнь, как сложный химический процесс. Опарин – как особую очень сложную форму движения материи. По определению Энгельса «жизнь - это способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей средой, причем с прекращением этого обмена прекращается и жизнь». Жизнь – это макромолекулярная открытая система, которой свойственна иерархическая организация, способность к самовозобновлению, обмен веществ и тонко регуляторный процесс. Основными свойствами живого являются сходный химический состав, обмен веществ, размножение, наследственность, изменчивость, рост, развитие, подвижность, раздражимость, приспособляемость. Ни одно из этих свойств не позволяет провести резкой границы между живым и неживым и лишь по совокупности этих особенностей можно охарактеризовать жизнь. Например, кристаллы способны к росту. Только живое обладает совокупностью всех перечисленных свойств. Более того, фундаментальные свойства живого - тесно связанные, неотделимые друг от друга феномены. Сходный химический состав (химический состав живого вещества представлен группой биогенных атомов ( N, Р, Са, К и др). Обмен веществом и энергией. Обмен веществ – особый способ взаимодействия живых организмов со средой. Обмен веществ требует постоянного притока некоторых веществ и энергии из вне и выделения некоторых продуктов диссимиляции во внешнюю среду. ДИССИМИЛЯЦИЯ (от лат. dis — отрицат. частица, assimilatio — употребление) — одна из двух составляющих процесса метаболизма, наряду с ассимиляцией. Диссимиляция — это совокупность процессов распада органических веществ с образованием более простых химических соединений (например, процесс гликолиза, расщепление жиров и др.). Эти процессы сопровождаются освобождением энергии и ресурсов, необходимых для осуществления синтетических процессов (например, биосинтеза белка или крахмала). Размножение, которое обеспечивает преемственность между поколениями организмов. Наследственность – поток информации между поколениями. Изменчивость – появление новых признаков в процессе репродукции.. Раздражимость – заключается в передаче информации от внешней среды к организму; на основе раздражимости осуществляется саморегуляция и гомеостаз. Жизнь на Земле представлена крайне многообразными формами. В зависимости от методов исследований во всем этом многообразии можно выделить различные уровни организации живого. Каждая более высокая форма материи включает в себя элементы более низкого уровня. Молекулярный уровень. Элементарные структурные единицы – молекулы. Основные элементарные явления этого уровня: репликация, биосинтез, мутации. Клеточный уровень. Структурными элементарными единицами являются различные органоиды и компоненты клеток. Основные элементарные явления этого уровня: способность к самовоспроизведению, регуляторность химических реакций, запасание и расходование энергии. Организменный уровень. Единицей уровня является организм. Основным процессом на данном уровне является онтогенез. Популяционно-видовой уровень. Популяция – элементарная структура на популяционно-видовом уровне, элементарное явление – изменение генотипического состава популяции. Основные признаки уровня: рождаемость, смертность, структура (половая и возрастная), плотность, численность популяции. Выделены элементарные факторы, действующие на этом уровне: мутационный процесс, популяционные волны, изоляция и естественный отбор. Биосферный уровень. Единицей уровня является биоценоз. Биоценоз – совокупность живых организмов, населяющих участок суши или водоема и находящихся в определенных отношениях между собой. Вместе с конкретными участками земной поверхности, занимаемыми биоценозами, и прилежащей атмосферой называются экосистемами. Экосистема – взаимообусловленный комплекс живых и косных компонентов, связанных между собой обменом веществ и энергией. Для этого уровня характерно: активное взаимодействие живого и неживого вещества, биологический круговорот веществ и энергии. Проблема происхождения жизни – одна из фундаментальных и наиболее сложных в современном естествознании. Существует множество гипотез происхождения жизни. Наиболее важными из них являются следующие: Креационизм. Самопроизвольное зарождение (самозарождение; жизнь возникала неоднократно из неживого вещества). Стационарного состояния (жизнь существовала всегда). Панспермия (жизнь занесена на Землю из других планет). Биохимические (жизнь возникла в условиях Земли в результате процессов, подчиняющихся химическим и физическим законам, т.е. в результате биохимической эволюции). Гипотезы самозарождения. На протяжении тысячелетий люди верили в самопроизвольное зарождение жизни, считая его обычным способом появления живых существ из неживой материи. Полагали, что источником спонтанного зарождения служат либо неорганические соединения, либо гниющие органические остатки (концепция абиогенеза). Эта гипотеза была распространена в древнем Китае, Вавилоне и Египте в качестве альтернативы креационизму. Идея самозарождения высказывали также философы Древней Греции. Постепенно эта гипотеза видоизменялась. Аристотель считал, что лягушки и насекомые заводятся в сырой почве. В средние века многим "удавалось" наблюдать зарождение насекомых, червей, угрей, мышей. Эти "факты" считались достаточно убедительными. В 1668 г. итальянский врач Франческо Реди опровергнул эти взгляды опытным путем. Для этого он поместил мертвых змей в разные сосуды, причем одни сосуды накрыл кисеей, а другие оставил открытыми. Налетевшие мухи отложили яйца на мертвых змей в открытых сосудах; вскоре из яиц вывелись личинки. В накрытых сосудах личинок не оказалось. Таким образом, Реди доказал, что белые черви, появляющиеся в мясе змей, являются личинками флорентийской мухи, и что если мясо закрыть и предотвратить доступ мух, то оно не "произведет" червей. Опровергнув концепцию самозарождения, Реди высказал мысль о том, что жизнь может возникнуть только из предшествующей жизни. Похожие опыты проделал итальянский ученый Ладзаро Спалланцани (1729-1799), который решил доказать опытным путем, что микроорганизмы, часто обнаруживаемые в мясном бульоне, самопроизвольно в нем не зарождаются. С этой целью он помещал жидкость, богатую органическими веществами (мясной бульон), в сосуды, кипятил эту жидкость на огне, после чего сосуды герметически запаивал. В итоге бульон в сосудах оставался чистым и свободным от микроорганизмов. Своими опытами Спалланцани доказал невозможность самопроизвольного зарождения микроорганизмов. Противники этой точки зрения утверждали, что самозарождение не происходит по той причине, что воздух в колбах во время кипячения портится, поэтому по-прежнему признавали гипотезу самозарождения. В XIX в. гипотеза самозарождения была окончательно опровергнута французским ученым Луи Пастером (1822-1895) и английским биологом Джоном Тиндалем. Они показали, что бактерии распространяются по воздуху и что если в воздухе, попадающем в колбы с простерилизованным бульоном, их нет, то и в самом бульоне они не возникнут. Пастер пользовался для этого колбами с изогнутым S-образным горлышком, которое служило для бактерий ловушкой, тогда как воздух свободно проникал в колбу и выходил из нее. Тиндаль стерилизовал воздух, поступающий в колбы, пропуская его сквозь пламя или пропуская через вату. К концу 70-х гг. XIX в. практически все ученые признали, что живые организмы происходят только от других живых организмов. Гипотеза стационарного состояния. Согласно этой гипотезе, Земля никогда не возникала, а существовала вечно; она всегда была способна поддерживать жизнь, а если и изменялась, то очень мало; виды также существовали всегда. Эту гипотезу называют иногда гипотезой этернизма (от лат. eternus – вечный). Гипотеза этернизма была выдвинута немецким ученым В. Прейером в 1880 г. Взгляды Прейера поддерживал академик В. И. Вернадский. Гипотеза панспермии (от греч. – весь, всякий и sperma– семя) –гипотеза о появлении жизнина Земле в результате переноса с других планет зародышей жизни. Эта гипотеза примыкает к гипотезе стационарного состояния. Ее приверженцы поддерживают мысль о вечном существовании жизни и выдвигают идею о внеземном происхождении. Одним из первых идею о космическом (внеземном) происхождении жизни высказал немецкий ученый Г. Рихтер в 1865 г. Согласно Рихтеру, жизнь на Земле не возникла из неорганических веществ, а была занесена из других планет. В связи с этим вставали вопросы, насколько возможно такое перенесение с одной планеты на другую и как это могло быть осуществлено. Сторонниками этой теории были Г. Гельмгольц, С. Аррениус, Дж. Томсон, П. П. Лазарев и др. Согласно представлениям Томсона и Гельмгольца, споры бактерий и других организмов могли быть занесены на Землю с метеоритами. Лабораторные исследования подтверждают высокую устойчивость живых организмов к неблагоприятным воздействиям, в частности к низким температурам. Например, некоторые споры и семена растений не погибают даже при длительном выдерживании в жидком кислороде или азоте. Другие ученые высказывали мысль о перенесении "спор жизни" на Землю светом. Современные приверженцы концепции панспермии (в числе которых лауреат Нобелевской премии английский биофизик Ф. Крик) считают, что жизнь на Землю занесена случайно или преднамеренно космическими пришельцами с помощью летательных аппаратов. Доказательством этого являются многократные появления НЛО (неопознанных летающих объектов), наскальные изображения предметов, похожих на ракеты и космонавтов, остатки сооружений, похожих на космодромы, а также сообщения о будто бы произошедших встречах с инопланетянами. К гипотезе панспермии примыкает точка зрения астрономов Ч. Вик-рамасингха (Шри-Ланка) и Ф. Хойла (Великобритания). Они считают, что в космическом пространстве, в основном в газовых и пылевых облаках, в большом количестве присутствуют микроорганизмы, где они, по мнению ученых, и образуются. Далее эти микроорганизмы захватываются кометами, которые затем, проходя вблизи планет, "сеют зародыши жизни". Биохимические гипотезы происхождения жизни. В настоящее время большинство биологов придерживаются теории происхождения биологических молекул абиогенным путем. В этой связи эта гипотеза будет рассмотрена отдельно. В основе современных научных представлений о происхождении жизни лежит гипотеза биохимической эволюции Опарина — Холдейна. Согласно теории биохимической эволюции формирование жизни на Земле шло в три этапа: абиогенный синтез органических веществ; образование биополимеров; формирование мембранных структур и появление самовоспроизведения. Абиогенный синтез органических веществ Согласно теории Опарина возникновение жизни на Земле возможно было только в условиях древней атмосферы и отсутствия живых организмов. На первых этапах своего существования наша Земля представляла собой раскалённый шар. По мере её остывания постепенно формировалась первичная атмосфера, состоящая из аммиака, метана, углекислого газа, цианистого водорода и паров воды. Ни кислорода, ни озона в атмосфере древней Земли не было. При дальнейшем понижении температуры образовался первичный океан. Под действием различных видов энергии (электрические разряды, ядерные реакции, солнечная радиация, извержения вулканов) образовались простые органические соединения: формальдегид, спирты, муравьиная кислота, аминокислоты и т. д. Окисление образовавшихся веществ не происходило, так как отсутствовал свободный кислород. Синтезированные вещества в течение десятков миллионов лет постепенно накапливались в древнем океане. Их накопление в итоге привело к образованию однородной массы — «первичного бульона». По мнению Опарина, именно в «первичном бульоне» и возникла жизнь. Этот этап биохимической эволюции был подтверждён экспериментально биохимиками С. Миллером, Дж. Оро и другими учёными.В экспериментальных установках, моделирующих условия первобытной Земли, ими были получены альдегиды, аминокислоты, простые сахара, пуриновые и пиримидиновые основания, нуклеотиды. Образование биополимеров Из простых органических веществ при определённых условиях синтезировались биополимеры. Аминокислоты соединялись в полипептиды, простые сахара превращались в полисахариды, а нуклеотиды — в нуклеиновые кислоты. Карбоновые кислоты, соединяясь со спиртами, могли образовать липиды, которые покрывали поверхность водоёмов жирной плёнкой. Возникшие белки формировали коллоидные комплексы, притягивающие к себе молекулы воды. Так появились коацерваты — сгустки органических веществ, обособленные от остальной массы воды. В коацерваты постоянно поступали органические соединения, в результате чего происходил синтез более сложных веществ. Они могли сливаться и увеличиваться в размерах.  Слияние коацерватных капель Образование биополимеров и коацерватов в условиях древней Земли подтверждено экспериментально работами Л. Орджела и С. Акабори. Ими были получены простейшие белки и нуклеотидные цепи. Формирование мембранных структур и появление самовоспроизведения Из липидных плёнок на поверхности коацерватов могла сформироваться биологическая мембрана. Объединение коацерватов с нуклеиновыми кислотами привело к образованию примитивных самовоспроизводящихся живых организмов — пробионтов. Эти первичные организмы были анаэробами и гетеротрофами и питались веществами «первичного бульона». Таким образом, около 3,5 млрд лет назад, согласно этой гипотезе, завершилось зарождение жизни на Земле. 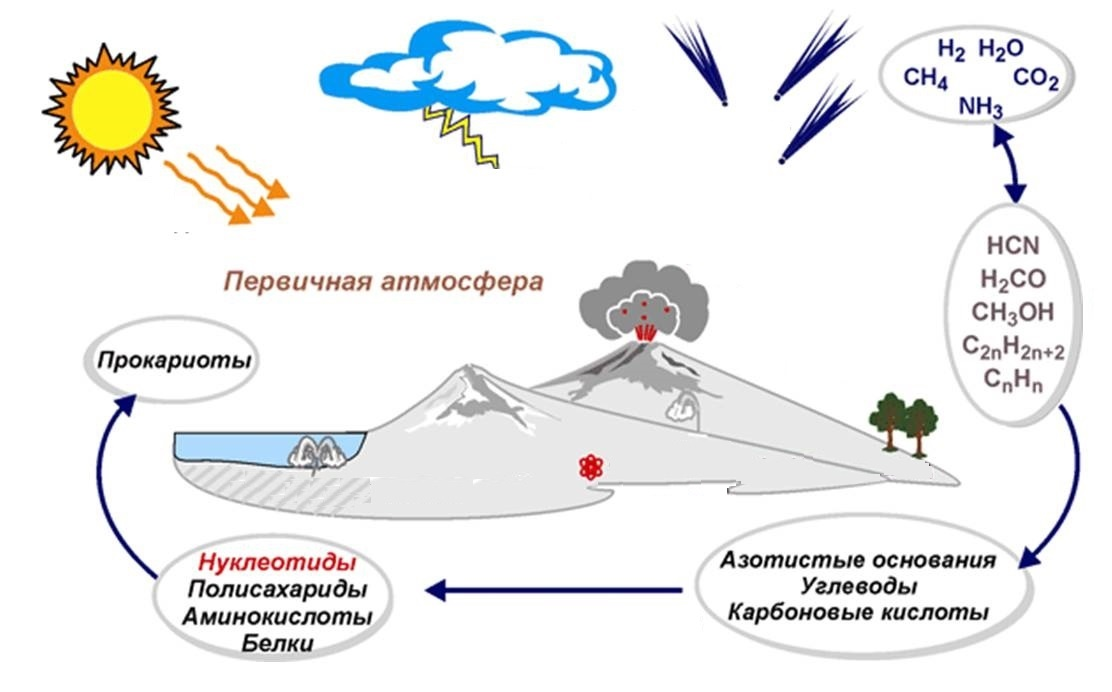 Краткие сведения о геохронологии Согласно современным оценкам возраст Земли составляет около 4,5-5 млрд. лет. Появление на планете первых водоемов, с которыми связывают зарождение жизни, отстоит от настоящего времени на 3,8-4 млрд. лет. Историю Земли принято делить на крупные промежутки времени – эры и периоды. Границами между ними являются крупные геологические события, связанные с историей развития планеты как космического тела. К таким событиям относятся горообразовательные процессы, усиление вулканической деятельности, поднятие и опускание суши, изменения очертаний материков и океанов. Геохронологическая история Земли состоит из 5 эр. Таблица 1. Хронология основных событий в эволюции многоклеточных организмов
Возникновение жизни. Основные этапы биогенеза В настоящее время большинство биологов придерживаются теории происхождения биологических молекул абиогенным путем. Эта теория была разработана независимо друг от друга русским биохимиком Александр Иванович Опарином (1924) и английским естествоиспытателем Джоном Холдейном (1929). Согласно этой теории, в условиях, имевших место на планете несколько миллиардов лет назад, было возможно образование живого вещества из неживого. К таким условиям относят: наличие атмосферы восстановительного типа; воды; источников энергии (в виде ультрафиолетового и космического излучения, теплоты остывающей земной коры, вулканической деятельности, атмосферных электрических явлений, радиоактивного распада); приемлемой температуры; отсутствие других живых существ. В 1953г. американский ученый Стенли Ллойд Миллер в условиях, иммитировавших атмосферу древней Земли, получил аминокислоты, жирные кислоты, мочевину, уксусную, муравьиную кислоты. Главные этапы на пути возникновения и развития жизни заключаются в следующем: образовании атмосферы из газов, которые могли бы служить «сырьем» для синтеза органических веществ (метана, оксида и диоксида углерода, аммиака, сероводорода, цианистых соединений), и паров воды; абиогенном образовании простых органических веществ (аминокислот, сахаров, азотистых оснований, АТФ и других мононуклеотидов); полимеризации мономеров в биологические полимеры (белки, нуклеиновые кислоты и др.); образовании предбиологических форм жизни (протобионтов), имеющих некоторые свойства живого; возникновении простейших живых форм (примитивных клеток), имеющих всю совокупность главных свойств жизни; биологической эволюции возникших живых существ. Возможность абиогенного образования органических веществв условиях, бывших на Земле около 4 млрд. лет назад, доказана опытами химиков. Кроме того, органические соединения обнаружены в космическом пространстве. В лабораторных условиях при пропускании электрических разрядов через различные газовые смеси, напоминающие примитивную атмосферу планеты, ученые получали аминокислоты (аланин, глицин, аспарагиновую кислоту), янтарную, уксусную, молочную кислоты, мочевину, азотистые основания (аденин, гуанин), АДФ и АТФ. Низкомолекулярные органические соединения накапливались в водах первичного океана в виде первичного бульона или же адсорбировались на поверхности глинистых отложений. По мере повышения концентрации полипептидов, полинуклеотидов и других органических соединений в первичном бульоне сложились условия для следующего этапа – самопроизвольного возникновения предбиологических форм сложного химического состава, или протобионтов. Предположительно они могли быть представлены коацерватами (А. И. Опарин) или микросферами (С. Фоке). Это коллоидные капли с уплотненным поверхностным слоем, имитирующим мембрану, содержимое которых составляли один или несколько видов биополимеров. Возможность образования в коллоидных растворах структур типа коацерватов или микросфер также доказана опытным путем. При определенных условиях протобионты проявляют некоторые общие свойства живых форм. Они способны в определенной степени избирательно поглощать и выделять вещества. Происходит процесс, напоминающий обмен веществ. Накапливая вещества, коацерваты увеличивают свой объем (рост). По достижении определенных размеров они распадаются на части, сохраняя при этом некоторые черты исходной химической организации (размножение). Поскольку устойчивость коацерватов различного химического состава различна, среди них происходит отбор. Протобионты представляются как обособленные от окружающей среды, открытые макромолекулярные системы, возникавшие в первичном бульоне и способные к примитивным формам роста, размножения, обмена веществ и предбиологическому химическому отбору. Предбиологическая эволюция протобионтов осуществлялась в трех главных направлениях. совершенствование каталитической функции белков. Так, включение железа в порфириновое кольцо гемоглобина увеличивает его каталитическую активность в сравнении с активностью самого железа в растворе в 1000 раз. приобретение протобионтами способности к самовоспроизведению. Это сделало возможным передачу информации от поколения к поколению. В основе этой способности лежит матричный синтез. Механизм матричного синтеза был использован также для переноса информации с полинуклеотидов на полипептиды. возникновение мембран. Вокруг коацерватов, богатых органическими соединениями, возникли слои липидов, отделивших коацерват от окружающей водной среды. Липиды преобразовались в ходе эволюции в наружную мембрану, существенно повысившую жизнеспособность и устойчивость организмов. Отграничение от окружающей среды мембраной с избирательной проницаемостью превращает протобионт в устойчивый набор макромолекул. Все это привело к возникновению живых организмов, которые поначалу были представлены примитивными безъядерными клетками (прокариотами). С момента появления клеток предбиологический химический отбор уступил место биологическому отбору. Дальнейшее развитие жизни шло согласно законам биологической эволюции. Эволюция одноклеточных организмов В течение длительного времени все организмы были гетеротрофными. Пищей им служили готовые органические вещества, накопленные в океане или другие гетеротрофные организмы. Постепенно запасы абиогенных органических веществ в первичном бульоне уменьшились. В связи с этим дальнейшее существование жизни оказалось под вопросом. Эта проблема разрешилась около 3,3-3,5 млрд. лет назад с развитием у организмов автотрофности. В связи с этим произошла дивергенция прокариот на два главных ствола их эволюции: бактерии (Bacteriae), в большинстве сохранившие в той или иной форме гетеротрофный способ питания; синезеленые водоросли (Cyanophyta), развившие автотрофное питание посредством фотосинтеза. Считается, что эукариотические организмы появились около 1,5 млрд. лет назад. Надцарство эукариот более миллиарда лет назад разделилось на царства животных, растений и грибов. Грибы более близки к животным, чем к растениям. По-видимому, многоклеточность возникла независимо у грибов, растений и животных. Важнейшие отличия эукариот от прокариот: дифференциация протоплазмы эукариотической клетки на отдельные органоиды (ядра, митохондрии и т. д.), имеющие различное строение и выполняющие различные функции; пространственная разобщенность процессов транскрипции и трансляции; локализация в ядре наследственного материала; более совершенный механизм клеточного деления (митоз, мейоз); возникновение цитоплазматической подвижности, обусловленное аппаратом микротрубочек. По поводу происхождения эукариотической клетки существует две основные теории. Это аутогенная теория (теория прямой филиации) и симбиотическая теория (теория последовательных эндосимбиозов). Аутогенная гипотеза. Согласно аутогенной гипотезе эукариотическая клетка возникала путем дифференциации исходной прокариотической клетки. Вначале образовалась мембрана, а затем из ее впячиваний возникли отдельные органоиды. Фотосинтезирующие эукариоты произошли от фотосинтезирующих прокариот. У образовавшихся автотрофных эукариотических клеток путем внутриклеточной дифференцировки образовались митохондрии. В результате утраты пластид от автотрофов произошли животные и грибы. Симбиотическая гипотеза. Эта теория является наиболее обоснованной и распространенной среди ученых. Впервые идею симбиогенеза выдвинул русский ученый А.С. Фоминцын, а детально ее разработала американская исследовательница Л. С. Маргулис. Согласно этой теории возникновение эукариотической клетки прошло через несколько этапов симбиоза с другими клетками. На первой стадии клетки типа анаэробных гетеротрофных бактерий включили в себя свободноживущие аэробные бактерии, превратившиеся в митохондрии. Параллельно этому в клетке-хозяине прокариотической генофор формируется в обособленное от цитоплазмы ядро. Таким путем возникла первая эукариотическая клетка, которая была гетеротрофной. Возникшие эукариотические клетки путем повторных симбиозов включили в себя синезеленые водоросли, что привело к появлению у них структур типа хлоропластов. Таким образом, митохондрии уже были у гетеротрофных эукариотических клеток, когда последние в результате симбиоза приобрели пластиды. В дальнейшем в результате естественного отбора митохондрии и хлоропласты утратили часть генетического материала и превратились в структуры с ограниченной автономией. Доказательства эндосимбиотической теории: Сходство структуры и энергетических процессов у бактерий и митохондрий, с одной стороны, и у синезеленых водорослей и хлоропластов, с другой стороны. Митохондрии и пластиды имеют собственную специфическую систему синтеза белков (ДНК, РНК, рибосомы). Специфичность этой системы заключается в автономности и резком отличии от таковой в клетке. ДНК митохондрий и пластид представляет собой небольшую циклическую или линейную молекулу, которая отличается от ДНК ядра и по своим характеристикам приближается к ДНК прокариотических клеток. Синтез ДНК митохондрий и пластид не зависит от синтеза ядерной ДНК. В митохондриях и хлоропластах имеются и-РНК, т-РНК, р-РНК. Рибосомы и р-РНК этих органоидов резко отличаются от таковых в цитоплазме. В частности рибосомы митохондрий и хлоропластов, в отличие от цитоплазматических рибосом, чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток. Увеличение числа митохондрий происходит путем роста и деления исходных митохондрий. Увеличение числа хлоропластов происходит через изменения пропластид, которые, в свою очередь, размножаются путем деления. Эта теория хорошо объясняет сохранение у митохондрий и пластид остатков систем репликации и позволяет построить последовательную филогению от прокариот к эукариотам. Возникновение и развитие многоклеточной организации Первые многоклеточные животные и растения появились на Земле в протерозое около 700-900 млн. лет назад. У растений возникновение многоклеточного уровня организации произошло на основе дифференциации лентообразных колоний. У колоний, прикрепленных одним концом к субстрату, различные участки находились в разных условиях по отношению к падающему свету, субстрату и водной среде. В связи с этим естественный отбор благоприятствовал возникновению некоторой дифференциации частей колонии. На одном конце колонии выделялись клетки, служившие для прикрепления к субстрату (для них характерно ослабление фотосинтеза, потеря способности к делению), на другом же конце - верхушечные клетки, интенсивно делившиеся и образовавшие своего рода "точку роста" колонии. Естественный отбор благоприятствовал приобретению клетками колонии способности делиться в разных направлениях; это приводило к ветвлению, что увеличивало поверхность колонии. Это вело к возникновению многослойного тела. В процессе дальнейшей дифференциации сформировались органы, выполнявшие разные функции (фиксация на субстрате, фотосинтез, размножение). Среди многочисленных гипотез о происхождении многоклеточности животных наиболее известны гипотезы немецкого зоолога Э.Геккеля и русского ученого И.И. Мечникова. Гипотеза Э. Геккеля основана на сформулированном им биогенетическом законе. В соответствии с этим Э.Геккель полагал, что филогенез древнейших Metazoa в определенной степени повторяется в онтогенезе современных низших многоклеточных животных. По Геккелю, предками Metazoa были колониальные простейшие, обладавшие сферическими колониями с однослойной стенкой, подобными бластуле. Геккель назвал эту гипотетическую предковую форму "бластеей". У бластеина переднем полюсе возникло впячивание ее стенки внутрь, подобно тому, как это происходит при гаструляции в онтогенезе некоторых современных Metazoa. Эту предковую форму за сходство с эмбриональной гаструлой дало основание Э.Геккелю назвать ее гастреей. Гастрея имела рот, слепо замкнутую кишечную полость и двуслойную стенку тела, состоящую из экто и энтодермы. Гастрея Геккеля близка по организации к кишечнополостным. И.И.Мечников обратил внимание на то, что у примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша - бластулы), что характерно для более высокоорганизованных групп, а посредством миграции некоторых клеток из однослойной стенки тела внутрь. Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. Такой способ гаструляции гораздо проще, чем инвагинация, так как не требует сложного направленного и координированного смещения целого пласта клеток, и, вероятно, примитивнее инвагинации. В связи с этим Мечников модифицировал гипотезу Геккеля следующим образом. В сфероидной колонии простейших - жгутиконосцев клетки ее однослойной стенки, захватывавшие пищу, мигрировали для ее переваривания внутрь, в полость колонии. Эти клетки образовали рыхлое внутреннее скопление - фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток - кинобласт - осуществлял функции защиты и движения организма. Для захвата новых пищевых частиц клеткам фагоцитобласта, по мысли Мечникова, не было необходимости возвращаться в поверхностный слой: располагаясь непосредственно под кинобластом, клетки фагоцитобласта захватывали пищевые частицы псевдоподиями, выдвигаемыми наружу в промежутках между клетками фагоцитобласта. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. В дальнейшем как приспособление к повышению активности питания у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. Гипотеза Мечникова, как и гипотеза Геккеля, рассматривает в качестве наиболее примитивных многоклеточных животных кишечнополостных и губок. Эволюция растительного мира Основные особенности эволюции растительного мира: постепенный переход к преобладанию диплоидного поколения над гаплоидным. У многих водорослей все клетки (кроме зиготы) гаплоидны, у голосеменных и покрытосеменных почти полностью редуцируется гаметофит и значительно удлиняется в жизненном цикле диплоидная фаза. независимое от влажной среды размножение. Переход от наружного оплодотворения к внутреннему, возникновение двойного оплодотворения и обеспечение зародыша запасами питательных веществ. дифференцировка наземных растений на корень, стебель и лист, развитие сосудистой проводящей системы, опорных и защитных тканей. совершенствование органов размножения и перекрестного опыления у цветковых в коэволюции с насекомыми. развитие зародышевого мешка для зашиты растительного эмбриона от неблагоприятных влияний внешней среды; возникновение разнообразных способов распространения семян и плодов физическими и биотическими факторами. Эволюция животного мира Эволюция животного мира изучена лучше, чем растений. Это связано с тем, что они обладают скелетом и поэтому лучше закрепляются в окаменелых остатках. Самые ранние следы животных обнаруживаются в конце докембрия (700 млн. лет). Предполагается, что первые животные произошли либо от общего ствола всех эукариот, либо от одной из групп древнейших водорослей. Наиболее близки к предкам простейших животных (Protozoa) одноклеточные зеленые водоросли. Так эвглену и вольвокс, способных и к фотосинтезу, и к гетеротрофному питанию, ботаники относят к типу зеленых водорослей, а зоологи к типу простейших животных. Наиболее существенные черты эволюции животного мира заключались в следующем: прогрессивное развитие многоклеточности и связанная с ним специализация тканей и всех систем органов. Свободный образ жизни (способность к перемещению) в значительной мере определил совершенствование форм поведения, а также автономизацию онтогенеза – относительную независимость индивидуального развития от колебаний факторов среды на основе развития внутренних регуляторных систем. возникновение твердого скелета: наружного - у членистоногих, внутреннего – у позвоночных. Такое разделение определило разные пути эволюции этих типов животных. Наружный скелет членистоногих препятствовал увеличению размеров тела, именно поэтому все насекомые представлены мелкими формами. Внутренний скелет позвоночных не ограничивал увеличение размеров тела, достигших максимальной величины у мезозойских рептилий – динозавров, ихтиозавров; развитие центральной нервной системы. Развитие центральной нервной специализации у насекомых характеризуется совершенствованием форм поведения по типу наследственного закрепления инстинктов. У позвоночных развился головной мозг и система условных рефлексов, наблюдается ярко выраженная тенденция к повышению средней выживаемости отдельных особей. Развитие социальности в ряде ветвей древа животных, финальным событием которого стало возникновение биосоциального существа – человека. |