рк ответы биохим. рк БХ2. 236 ом бх рк2 атымтай аружан, жаксылыкова айсауле, нуртулеу жансая, шымкентбай данагуль
 Скачать 1.07 Mb. Скачать 1.07 Mb.
|

|
5 кофермент қатысады: Тиаминдифосфат (ТДФ) Флавинадениндинуклеотид (ФАД) Ацетил-КоА немесе кофермент А Никотинамидадениндинуклеотид (НАД) Липой қышқылы және оның амиді Пирожүзім қышқылының тотыға декарбоксилденуі 5 кезеңнен тұрады: 1-кезең Пируват Е1 (Пируватдегидрогеназа) құрамындағы ТДФ-пен әрекеттесіп, декарбоксилденеді, яғни карбоксил тобынан СО2 бөлініп шығады. Нәтижесінде гидрокси этил түзіледі. 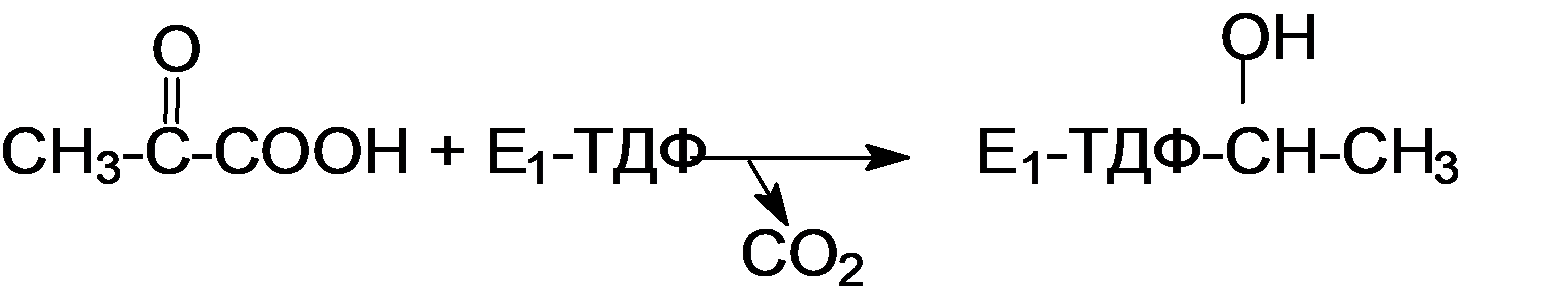 2-кезең Е2 (Дигидролипоилацетилтрансфераза) сутек атомдарын және ацетил тобын ТДФ-тан липоамидке жеткізеді. Нәтижесінде липой қышқылының ацетил тиоэфирі түзіледі.  3-кезең Ацетил КоА Е2 (Дигидролипоилацетилтрансфераза)-нің ацетил туындысымен әрекеттеседі. Нәтижесінде ацетил КоА-ның және липоамидтің тотықсызданған түрі түзіледі. 4-кезең Е3 (Дигидролипоилдегидрогеназа) сутек атомдарын тотықсызданған липоамидтен ФАД-қа тасымалдайды. Нәтижесінде, тотықсызданған ФАД түзіледі. 5-кезең Тотықсызданған ФАД сутек атомдарын НАД-қа жеткізеді. Нәтижесінде НАДН түзіледі. Қорытынды: пируваттың тотыға декарбоксилденуі кезінде, 1 моль пируваттан 3 моль АТФ түзіледі. Лимон қышқылының циклі Басқаша цитратты цикл, Кребс циклы, үш карбон қышқыладры циклы д.а. Пирожүзiм қышқылының тотыға декарбоксилденуi нәтижесiнде түзiлген Ацетил-КоА үш карбон қышқылдарының циклiне қатысады. Катаболизмнің қорытынды кезеңі, яғни бұл кезеңде ацетил- КоА-ның ацетил тобының көміртек атомдары 2 молекула СО2-на дейін тотығады. 8 реакциядан тұрады. 1-кезең-Цитраттың түзілуі Ацител-Ко-А қымыздық сірке қышқылымен, яғни оксалоацетатпен байланысып, цитрат және коэнзим А бөлініп шығады. 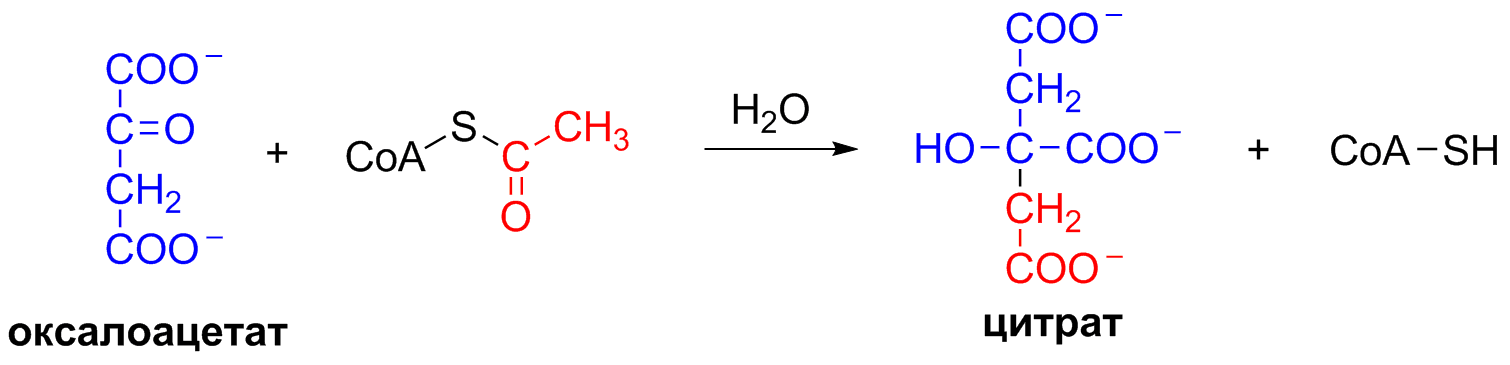 2-кезең-Цитраттың изоцитратқа айналуы Бұл реакцияны катализдейтін фермент- аконитаза. Ол суды цис-аконитаттың қос байланысына қосады. Нәтижесінде, біреуінен цитрат, ал екіншісінен изоцитрат түзіледі. Сондықтан да, бұл реакция қайтымды реакция болып табылады. 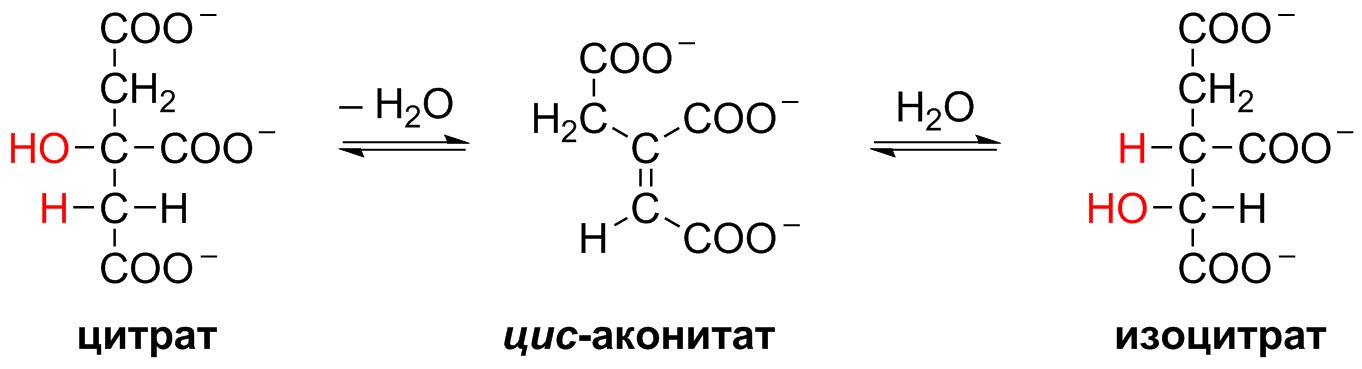 3-кезең-Изоцитраттың тотыға декарбоксилдену реакциясы Бұл реакцияны изоцитратдегидрогеназа ферменті катализдейді. Ферменттің 2 изоферменттік түрі бар: бірінің коферменті NAD+. Екіншісінің коферменті NADP+. Ферменттің әсері нәтижесінде изоцитраттан α-кетоглутарат түзілед. 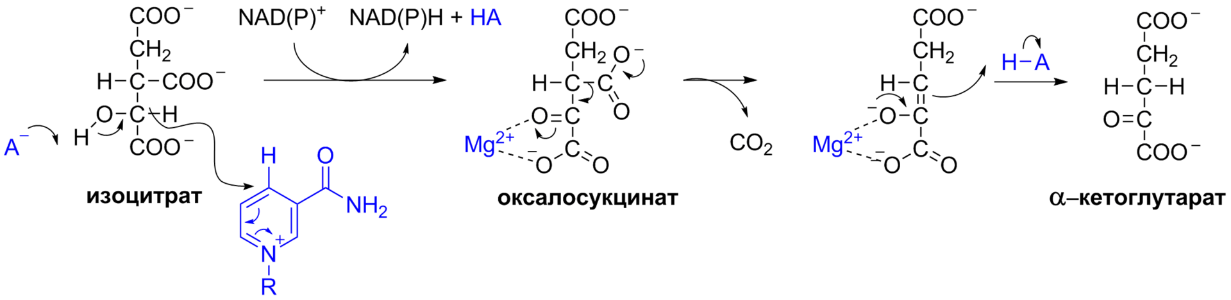 4-кезең-α-кетоглутараттың тотыға декарбоксилденуі Реакцияны α-кетоглутаратдегидрогеназа ферменті катализдейді. Бұл кезде NAD+ электрон акцепторы, ал КоА сукциния тобының тасымалдаушысы ретінде қызмет етеді. 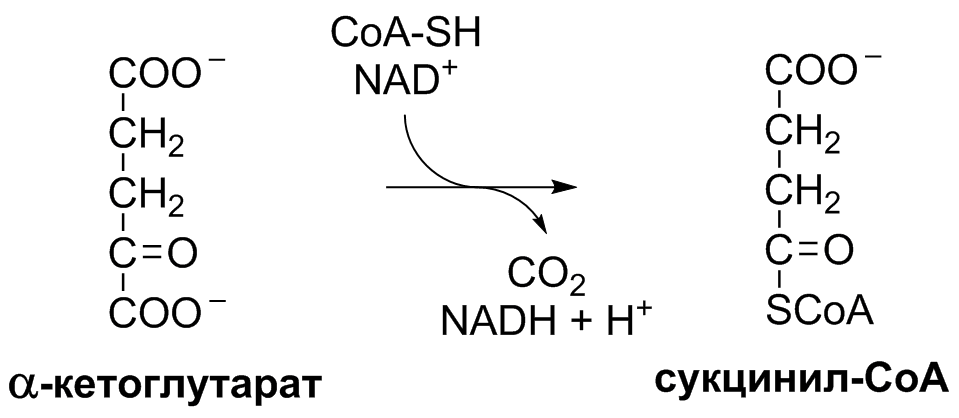 5-кезең-Сукцинил-КоА сукцинатқа айналуы Бұл қабысу реакциясын сукцинил КоА синтетаза ферментікатализдейді. Реакция кезінде бөлінетін энергия АТФ немесе ГТФ фосфоангидридтік байланыс түзуге жұмсалады. 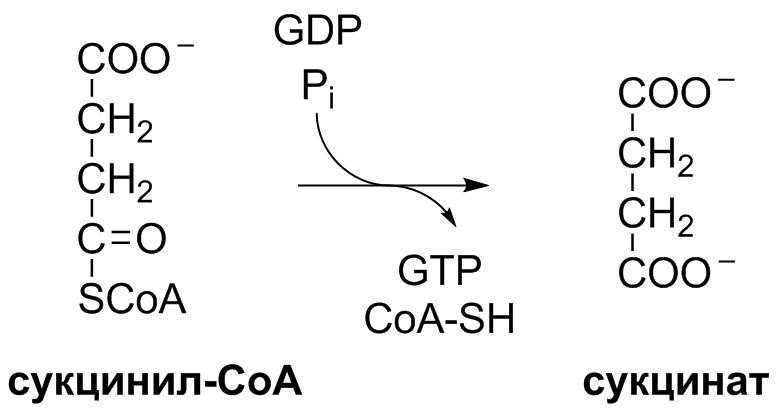 6-кезең-Сукцинаттың дегидрленуі Сукцинат сукцинат дегидргеназа ферментініңәсерінен фумаратқа айналады. 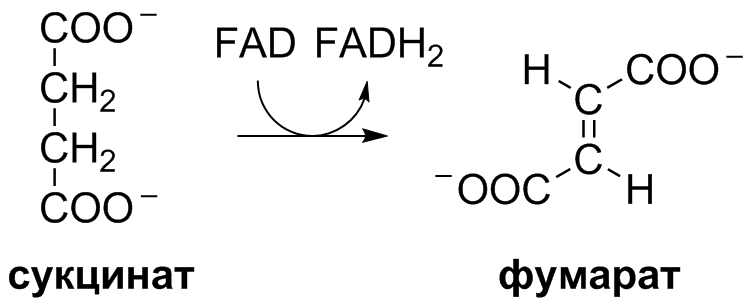 7-кезең-Фумараттан малаттың түзілуі Малаттың түзілуі фумарат гидратаза ферментінің қатысуымен жүреді. Бұл фермент көбінесе фумараза деген атпен белгілі. 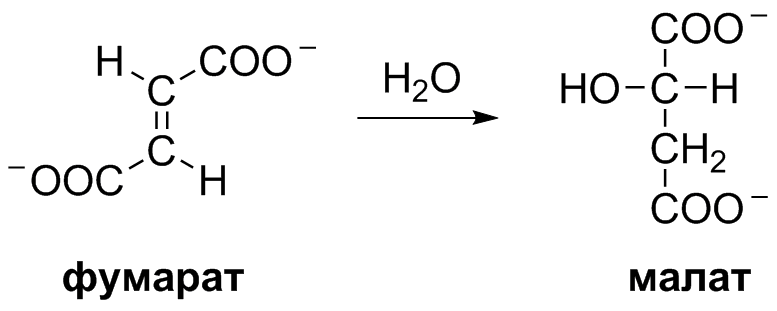 8-кезең-Малаттың дегидрленуі Цитратты циклдің аяқталу кезеңінде малат қымыздық сірке қышқылын түзе дегидрленеді. Реакцияны NAD –қа тәуелді малатдегидрогеназа ферменті катализдейді. 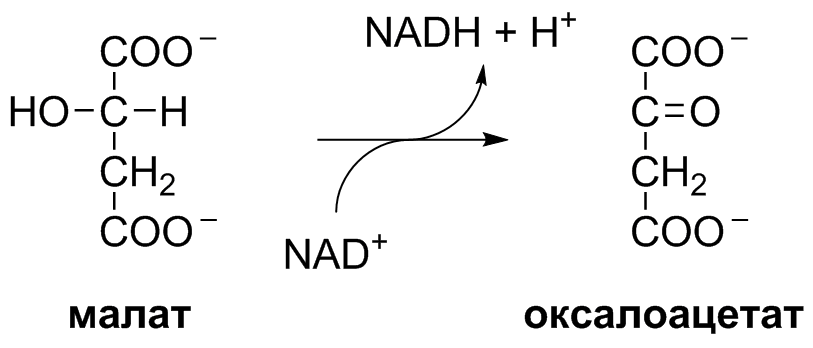 Қорыта келгенде, лимон қышқылының әрбір айналымында тотыға фосфорлану нәтижесінде 11 молекула АТФ-түзіледі. Бір молекула АТФ субстратты фосфорлану нәтижесінде түзіледі. !!! ТАТ пен катаболизмнің жалпы жолдарының жалпы реттелуі не нашла. 9.Ферменттердің митохондриялық ТАТ жинақталуы. Тынысалуға бақылау. Тыныс алу тізбегі митохондрияда орналасқан тыныс алу ферменттерінің комплекстерінен тұрады. Тыныс алу тізбегінің құрамы: Оксидоредуктаза классының ферменттері жатады. Бұл ферменттің ақуыздық емес бөлімін, яғни коферменті ролін РР дәруменнің туындысы НАД атқарады. Тотықсызданған НАДН*Н электрондар мен протондар флавопротеидтер немесе флавинді фермент, НАДН дегидрогеназа қабылдайды. Бұл ферменттің коферменті В2 дәруменінің туындысы-ФМН немесе ФАД болып табылады. Тыныс алу тізбегінің келесі звеносы убихинон ферменті. Ол майда еритін дәрумен, мембрананың липидтік қабатынан еркін өте алады, ақуызбен байланысқан немесе байланыспаған күйде кездеседі. Убихиноннан оттегіне электрондар цитохромдар арқылы тасымалданады. Цитохромдар-гем тәрізді құрылымнан тұратын күрделі ақуыздар. Митохондриядағы тыныс алу қарқынының АДФ концентрациясына тәуелділігін тыныс алуды бақылау деп аталады. Тыныс алуды бақылау механизмі өте жоғары дәлдігімен сипатталады және өте маңызды. Себебі, оның әсер ету нәтижесінде АТФ синтезінің жылдамдығы жасушаның энергияға мұқтаждығына сәйкес келеді. Жасушада АТФ қоры болмайды. АТФ/АДФ салыстырмалы концентрациясы ұлпаларда өте аз мөлшерде өзгереді, ал жасушаның энергияға мұқтаждығы, яғни АТФ ондаған есеге өзгеруі мүмкін. Организмдегі АТФ мөлшері 30-50 г, бірақ АТФ-тың рәбір молекуласы жасушада бір минуттан аз өмір сүре алады. Тәудігіне, адам организмінде 40-60 кг-ға дейін АТФ синтезделіп, сонша АТФ ыдырайды. АДФ концентрациясының артуы бірден тыныс алуды және фосфорлануды жылдамдатады. 10.Фосфорлана тотығу – АТФ синтезінің негізгі механизмі. Митчельдің хемосмостық теориясы. Фосфорлану коэффиценті. Тотығу фосфорлану- бұл тірі жүйенің ішінде жасушалық АТФ-тің көп бөлігін шығаратын ең негізгі механизм. Тотыға фосфорлану АТФ синтезі кезінде молекулалық оттегін қолданады және ол митохондрияның мембраналарына жақын жерде жүреді. Тотыға фосфорлану дегеніміз - оттегі қатысуымен ферменттердің көмегімен АТФ түзетін метаболикалық жол. Бұл аэробты организмдердің жасушалық тыныс алуының соңғы сатысы. Тотыға фосфорлануының екі негізгі процесі бар: электронды тасымалдау тізбегі химиозмоз Электронды тасымалдау тізбегінде ол электрондар донорларынан электронды акцепторларға электрондардың қозғалысын қоздыруға көптеген тотықтырғыш аралықтары қатысатын тотығу реакциясын жеңілдетеді. Осы тотығу реакцияларынан алынған энергия химиозмозда АТФ түзуге жұмсалады. Эукариоттар жағдайында тотығу фосфорлануы митохондрияның ішкі мембранасында әртүрлі ақуыздық кешендерде жүреді. Прокариоттар аясында бұл ферменттер клетканың аралық кеңістігінде болады. Тотығу фосфорлануына қатысатын белоктар бір-бірімен байланысады. Эукариоттарда электронды тасымалдау тізбегі кезінде бес негізгі ақуыз кешені қолданылады. Тотығу фосфорлануының соңғы электронды акцепторы оттегі болып табылады. Ол электронды қабылдайды және су түзілуіне дейін азаяды. Демек, тотығу фосфорлануымен АТФ түзілу үшін оттегі болуы керек. Тотыға фосфорлану коэффициенті Тотыға фосфорлану коэффициенті-АДФ-тың фосфорлануына жұмсалған фосфор қышқылы (Р) санының, тыныс алу кезінде сіңірілген оттек атомы санына қатынасы. Ол Р/О деп белгіленеді. Соған сәйкес НАДН үшін Р/О=3, ал сукцинат үшін Р/О=2-ге тең. Көрсетілген мәндер АТФ-синтезінің теория жүзінде максимум мәнін көрсетеді, ал іс жүзінде төмен болып келеді. Қазіргі кезде кең таралған П.Митчелл ұсынған хемиосмостық гипотеза болды. Митчелдің айтуы бойынша: Тыныс алу тізбегі бойынша электрондар тасымалданған кезде протондар матрикстен мембрана-аралық кеңістікке жеткізіледі. Осының нәтижесінде мембрананың екі жағында протон иондарының коцентарция градиентінде айырмашылық туады. Протондардың мембрана-аралық кеңістікке өтуі мембрана сыртын оң зарядтап, ішін теріс зарядтайды. Нәтижесінде трансмембраналық потенциал пайда болады. Осы екі потенциал протондық потенциалды құрайды. Сутек иондары АТФ синтетаза ферменті молекуласындағы арнаулы каналдар арқылы қайтадан матрикске өтуге тырысады. Міне, осы протондардың қайта өтуі кезінде бөлінген энергия тыныс алумен фосфорланудың қатар жүруінің қозғаушы күші болып табылады. Міне, Митчелл гипотезасының негізгі мағынасы осында. 11. Оттегінің улы түрлері, оларды залалсыздандыру ферменттері, антиоксиданттар. Жасушаға түскен О2 90% сіңіріледі. Ал қалған бөлігі басқа тотығу-тотықсыздану реакцияларында қолданылады. Оттекті қолдана отырып, тотығу-тотықсыздану реакцияларына қатысатын ферменттер, 2 топқа бөлінеді: Оксидазалар-оттекті тек электрондар акцепторы ретінде қолданып, оны Н2О немесе Н2О2 (сутегі тотығы) дейін тотықтырады. Оксигеназалар-бір немесе екі оттек атомын реакция нәтижесінде түзілетін өнімге енгізеді. Бұл реакциялар АТФ синтезделуін жүргізбеседе, олар көптеген арнайы реакцяилар амин қышқылдары алмасуында, өт қышқылдары және стероидтар синтезі, бауырда бөгде заттарды залалсыздандыру үшін қажет. Оттектің қатысуымен жүретін көптеген реакцяиларда, оның тотықсыздануы біртіндеп, әрбір кезең сайын бір электронды тасымалдау арқылы жүреді. Бір электронды тасымалдау кезінде оттектің реакцияға т.су қабілеті жоғары аралық заты түзіледі. Қозба,ан жағдайда оттектің улылығы жоқ. Оттектің улы түрлерінің түзілуі, оның молекулалық құрылысының ерекшеліктерімен байланысты. Оттектің белсенді түрлері, көптеген қосылыстардан электрондарды бөліп алады, нәтижесінде олар жаңа бос радикалдарға айналады, тізбекті тотығу реакциялары күшейеді. Оттектің белсенді түрлері әсерінен организмді барлық жасушаларда болатын, талғамдақтары жоғары ферменттер: супероксиддисмутазалар, каталазалар, глутатионпероксидазалар, сонымен қатар антиоксиданттар қорғап отырады. Антиоксиданттар-тотығуға қарсы және оны тежеу үшін қолданылатын заттар, ағзаның қартаюын бәсендететін қышқылдану процестерін баяулататын химиялық заттар тобы. Олар ағза үшін зиянды, ағзаға бұзушы әсер ететін бос радикалдарды бейтараптандырады. 12. Ұлпалық тыныс алудың бұзылуына және катаболизмнің жалпы жолдарына байланысты патология. Не нашла 13.Ұлпалық тыныс алудағы ингибиторлар. Фосфорлана тотығу мен ұлпалық тыныс алу процесінің ажыратушылары. Электрондарды тасымалдау тізбегін зерттеу, ұлпалық тыныс алу процесінің белгілі бір кезеңдеріне бөгет болушы арнайы тежеушілердің әсерін зерттеуге мүмкіндік береді. Тежелген кезеңнің алдында тікелей тұрған электронды тасымалдаушылардың тотықсыздану дәрежесі артса, ал одан кейінгі тұрған тасымалдаушының тотығу дәрежесі артады. ТАТ ингибиторлардың әсер ету орны: НАДН-дегидрогеназаның ингибиторлары: ротенонулылығы өте күшті зат кейбір балдырларда кездеседі, олар балықтар үшін у болып табылады, амиталбарбитуаттар тобына жататын дәрілік препарат. QH2-дегидрогеназаның ингибиторы-А антимицині, ол улы антибиотик болып табылады. Цитохромоксидазаның ингибиторлары цианид. Ол адам үшін өте улы және ол цитохромоксидазада электрондардың оттекке тасымалдануына бөгет болады. Кейбір химиялық заттар протондарды және басқа иондарды мембрана аралық кеңістіктен АТФ-синтаза каналы арқылы емес, мембрана арқылы матрикске тасымалдай алады. Нәтижесінде, электрохимиялық потенциал жойылады және АТФ синтезі тоқталады. Бұл құбылыс тыныс алу мен фосфорланудың ажырауы деп атайды. Ажырау нәтижесінде АТФ төмендейді, ал АДФ мөлшері артады. Ажыратушылар-липофильді заттар, яғни мембрананың липидтік қос қабатынан оңай өтетін заттар. Осындай заттың бірі-2,4-динитрофенол, яғни ионданған күйде мембрана аралық кеңістіте протонды қосып алып ионданбаған күйде матрикске өтіп, протондарды матрикске жеткізеді. Ажыратуышлар ролін, сонымен қатар дәрілік препараттар атқарады. Мысалы, дикумарол-антикоагулянт және организмде түзілетін метаболиттер, билирубин-гем катаболизмінің өнімі, тироксин-қалқанша безі гормоны. 14. Биомембрананың функциясы мен құрылысы және зат алмасудағы ролі. Барлық жасушаларда мембраналар бар. Прокариоттарда-цитоплазмалық мембрана болса, эукариоттарда-цитоплазмалық мембрана, митохондрияның ішкі және сыртқы мембранасы, лизосомада, рибосомада өзінің мембранасы болады, яғни клеткада әртүрлі мембраналар кездеседі. Мембрананың функциясы: Жасушаны қоршаған ортадан бөліп тұру және жасуша ішілік компартменттерді қалыптастырады. Мембрана арқылы көптеген заттардың өтуін реттеу және бақылау. Жасуша аралық әрекеттерді қамтамасыз етеді және жасуша ішіне сигналдарды өткізуге қатысады. Тамақтың органикалық заттар энергиясын, АТФ молекласы химиялық байланыстар энергиясына түрлендіреді. Мембрананың құрылысы: Мембраналар липид пен ақуыздардан тұрады. Майлардың ақуыздармен салмақтық қатынасы 1:4 немесе 4:1 болып келеді. Сулы ортада майлардың молекулалары өте жылдам липидтік қосқабат түзеді, ал мұндай қосқабаттан полярлы молекулалар өте алмайды. Мембраналардың құрамына майлардың негізгі үш тобы кіреді: фосфолипидтер, сфинголипидтер, гликолипидтер. Мембрананың көптеген қызметі оның құрамндағы ақуыздардың қасиетімен анықталады. Мембрана ақуыздары насостық, рецепторлық, ферменттік қызмет атқарады. Мембрана құрамындағы ақуыздардың 2 түрі болады: Шеткейлік ақуыздар-липидтермен электростатикалық күштер арқылы және сутектік байланыстардың көмегімен байланысып тұрады. Интегралды ақуыздар-мембраналық липидтердің көмірсутектік тізбектерімен көптеген байланыс түрлері арқылы байланысатындықтан, оларды бөліп алу қиын. Оларды бөліп алу үшін арнайы детергенттер, агенттер қолданылады. Мембрананың зат алмасудағы ролі. Мембрана арқылы заттардың алмасуының 3 жолы бар: Қарапайым диффузия-О2, СО2, Н2О және ауруды басушы кейбір дәрі-дәрімектер. Бұл заттардың өту жылдамдығы олардың мембранада ерігіщтігіне, диффузиялық кэффициентіне және осы заттардың клетка ішіндегі, сыртындағы концентрациясына тәуелді. Жеңілдетілген диффузия-мембрана арқылы өтетін заттың концентрациясы артқан сайын олардың өту жылдамдығы белгілі бір максимумге ұмтылады. Жеңілдетілген диффузияда өтетін зат тасымалдаушы бір затпен қосылып, клетка ішіне өтеді, онан соң ақуыз өзі өткізген заттан босап, қайтадан сыртқа шығады. Белсенді тасымалдау. Бұл кезде өтетін зат коцентрациялық градиентке тәуелді емес. Бұл үшін энергия қажет. Белсенді тасымалдау-белсенді тасымалдаушы АТФ-азалардың немесе ионды насостардың қатысуымен АТФ энергиясы есебінен іске асады. Ионды насостар-бұл АТФ-ты гидролизге және тасымалдаушы ионды талғамды қосып алуға қабілетті ақуызды құрылмдар. 15. Заттардың мембрана арқылы тасымалдану механизмі. Мембрана арқылы заттардың алмасуының 3 жолы бар: Қарапайым диффузия-О2, СО2, Н2О және ауруды басушы кейбір дәрі-дәрімектер. Бұл заттардың өту жылдамдығы олардың мембранада ерігіщтігіне, диффузиялық кэффициентіне және осы заттардың клетка ішіндегі, сыртындағы концентрациясына тәуелді. Егер, заттар коцентарциясының трансмембраналық градиенті болса, онда концентрация төмен жағына өту жылдамдығы жоғары болады. Концентрация екі жақты теңелгенше, заттардың тасымалдануы жүре береді. Жеңілдетілген диффузия- мұнда молекулалар концентрациясы төмен жаққа қарай өтеді. Мембрана арқылы өтетін заттың концентрациясы артқан сайын олардың өту жылдамдығы белгілі бір максимумге ұмтылады. Жеңілдетілген диффузияда өтетін зат тасымалдаушы бір затпен қосылып, клетка ішіне өтеді, онан соң ақуыз өзі өткізген заттан босап, қайтадан сыртқа шығады. Жеңілдетілген диффузия кезінде заттар мембрана арқылы тасымалдаушы-мембраналық ақуыздар көмегімен тасымалданады. Белсенді тасымалдау. Бұл кезде өтетін зат коцентрациялық градиентке тәуелді емес. Бұл үшін энергия қажет. Белсенді тасымалдау-белсенді тасымалдаушы АТФ-азалардың немесе ионды насостардың қатысуымен АТФ энергиясы есебінен іске асады. Ионды насостар-бұл АТФ-ты гидролизге және тасымалдаушы ионды талғамды қосып алуға қабілетті ақуызды құрылмдар. |