ботаника. 8. Типы пластид. Строение и специфические функции пластид, размножение и взаимопревращение
 Скачать 7.57 Mb. Скачать 7.57 Mb.
|

1 2 8. Типы пластид. Строение и специфические функции пластид, размножение и взаимопревращение. Пластиды (от греч. пластос - вылепленный, созданный и идос - вид) - мембранный органоид эукариот, относящихся к царству растений. Их строение напоминает строение митохондрий, так как пластиды тоже сформировались в процессе симбиоза. Очевидно, их эволюционными предками были бактерии, которые приобрели способность использовать световую энергию и были "захвачены" другими видами бактерий. Пластиды могут размножаться, имеют наружную и внутреннюю мембраны, межмембранное пространство и строму (от греч. строма - подстилка), соответствующую матриксу митохондрий (рис.). В строме имеются рибосомы бактериального типа и кольцевая молекула ДНК. В отличие от митохондрий, пластиды содержат в строме замкнутые мембранные образования - тилакоиды (от греч. тиллис - мешок и идос - вид). Пластиды — это органоиды, свойственные клеткам фотосинтезирующих эукариот — растений и водорослей. В зависимости от особенностей строения, окраски и выполняемых функций у растений выделяют три основных типа пластид: хлоропласты, лейкопласты и хромопласты. Внутреннее содержимое пластид называется стромой. Она окружена двумя мембранами. Между наружной и внутренней мембранами имеется межмембранное пространство. Лейкопласты - неокрашенные пластиды, как правило, выполняют запасающую функцию. В лейкопластах клубней картофеля накапливается крахмал. Лейкопласты высших растений могут превращаться в хлоропласты или хромопласты. Хромопласты - пластиды, окрашенные в жёлтый, красный или оранжевый цвет. Окраска хромопластов связана с накоплением в них каротиноидов. Хромопласты определяют окраску осенних листьев, лепестков цветов, корнеплодов, созревших плодов. Хлоропласты - пластиды, несущие фотосинтезирующие пигменты - хлорофиллы. Имеют зелёную окраску у высших растений, харовых и зелёных водорослей. Набор пигментов, участвующих в фотосинтезе (и, соответственно, определяющих окраску хлоропласта) различен у представителей разных таксономических отделов. Хлоропласты имеют сложную внутреннюю структуру.  Рисунок 1. Строение пластид: 1 – наружная мембрана, 2 –внутренняя мембрана, 3 – матрикс (строма), 4 – ламеллы стромы, 5 – грана, 6 – тилакоид, 7 – крахмальное зерно, 8 – липидная капля с пигментами. Все виды пластид образуются из мелких бесцветных пропластид (от лат. про - раньше, до), округлых мембранных телец диаметром около 1 мкм, которые имеют слабо развитую систему тилакоидов. Лейкопласты (от греч. лейкос - белый) - это бесцветные пластиды с немногочисленными тилакоидами, в с троме которых запасаются питательные вещества (крахмал, белки, липиды). Хромопласты (от греч. хрома - цвет, окраска) представляют собой окрашенные пластиды с отдельными или отсутствующими тилакоидами, содержащие каротиноиды (от лат. карота - морковь и греч. идос - вид, подобие) - желтые, оранжевые и красные пигменты (от лат. пигментум - краска). Они образуются в различных органах растений, придавая им специфическую окраску, которая привлекает животных, способствуя опылению цветков и распространению семян. Хлоропласты (от греч. хлорос - зеленый) - это пластиды зеленого цвета, что обусловлено наличием в них особого магнийсодержащего пигмента хлорофилла (от греч. филлон - лист), для образования которого необходим свет. Этот вид пластид описан немецким ботаником А. Шимпером в 1883 г. У водорослей они называются хроматофорами (от греч. хрома - цвет, окраска и форос - несущий). За многолетние исследования строения хлорофилла и других растительных пигментов немецкий химик Р. Вильштеттер в 1915 г. был отмечен Нобелевской премией. Кроме хлорофилла, хлоропласты содержат и каротиноиды, что объясняет осеннюю окраску листьев, потерявших способность синтезировать хлорофилл. Хлоропласты имеют форму двояковыпуклой линзы размером 4-6 мкм. В клетках высших растений их число может составлять от 20 до 50. Хлоропласты имеют наиболее совершенную организацию тилакоидной системы - отдельные тилакоиды собраны в стопки-граны (от лат. гранум - крупинка), которые соединены межгранными тилакоидами. Важнейшей функцией хлоропластов является синтез простых органических соединений из углекислого газа и воды. Исходная энергия, необходимая для этого, - видимый свет, поэтому данный процесс называют фотосинтезом (от греч. фотос - свет и синтезис - соединение, составление). Хлоропласты, хромопласты и лейкопласты способны к взаимному переходу. Так, при созревании плодов или изменении окраски листьев осенью хлоропласты превращаются в хромопласты, а лейкопласты могут превращаться в хлоропласты, например, при позеленении клубней картофеля. Пластиды поддерживают жизнедеятельность автотрофных клеток растений. Три вида органоидоидов отвечают за свои процессы, четко «делят обязанности», а в случае неблагоприятных условий трансформируются в необходимый для выживания органоид. 23. Типы основных тканей: фотосинтезирующая, запасающая, воздухоносная и водопоглощающая. Расположение в органах, строение, функции. Группы клеток, сходные по строению, происхождению и выполняемым функциям, образуют ткани. Выделяют следующие типы тканей: фотосинтезирующая, запасающая, воздухоносная и водопоглощающая. Фотосинтезирующая ткань. Ткани, основной функцией которых является работа ассимиляции, то есть фотосинтез, объединяют в систему ассимиляционных тканей. У высших растений они имеют обычно зеленую окраску. Поэтому могут быть названы зеленой паренхимой или хлорофиллоносной паренхимой (хлоренхимой). Ассимиляционная ткань устроена достаточно просто и состоит из однородных тонкостенных клеток. Хлоропласты в клетках хлоренхимы обычно расположены в один ряд в постенном слое цитоплазмы. Центральная часть полости клетки занята крупной вакуолью. Доступ углекислоты к клеткам хлоренхимы облегчается тем, что в ней имеется развитая система межклетников, сообщающаяся с атмосферой. Наличие межклетников является характернейшей особенностью ассимиляционной ткани. Межклетники обеспечивают газообмен с окружающей средой. В соответствии с тем, что работа ассимиляции происходит за счет солнечной энергии, хлоренхима располагается в местах, наиболее доступных свету: она находится в надземной части растений непосредственно под кожицей листьев и стеблей. Нередко хлоренхима дифференцирована на столбчатую (палисадную) и губчатую ткань. Палисадная ткань обычно состоит из удлиненных клеток цилиндрической формы, расположенных перпендикулярно к поверхности органа. Межклеточники в палисадной ткани развиты слабо. Палисадная ткань содержит большое количество хлоропластов, здесь происходят световые реакции фотосинтеза. Губчатая ткань построена из округлых или неопределенной формы клеток, образующих рыхлую сложную сетчатую систему. Межклетники хорошо развиты. Здесь протекает газообмен и темновая стадия фотосинтеза. 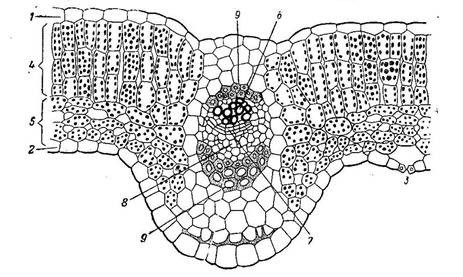 Рисунок 2. Часть поперечного разреза листа редьки: 1- верхняя кожица, 2- нижняя кожица, 3устьице, 4- палисадная паренхима, 5- губчатая паренхима. В середине сосудо-волокнистый пучок: 6- ксилема его, 7-камбий, 8-флоэма, 9- склеренхимная обложка (из Раздорский В.Ф., 1949) Запасающие ткани Поглощенные растением синтезированные вещества могут откладываться в виде запасов. К накоплению запасных веществ в той или иной мере способны все живые клетки, но в том случае, когда запасающая функция гипертрофированна (выступает на первое место), говорят о запасающих тканях. Запасы могут храниться длительное время, например, зимние запасы крахмала в корнях и клубнях двулетних и многолетних растений. В других случаях запасы почти непрерывно потребляются и вновь пополняются в период вегетации. В строении запасающих клеток встречается большое разнообразие.  Рисунок 3. Запасающая ткань в эндосперме пшеницы: 1 – алейроновый слой, 2 – ткань, содержащая крахмал (по В.Г.Александрову) Некоторые запасающие ткани приспособлены к накоплению воды. Их называют водоносными. Как правило, клетки водоносных тканей чрезвычайно гигроскопичны (т.е. способны быстро впитывать влагу) и имеют тонкие оболочки. Наличие водоносных тканей характерно для некоторых ксерофитов ≈ растений, приспособленных переносить длительный засушливый период. Чаще всего водоносные ткани встречаются в листьях. Они являются резервуарами влаги. При подсыхании растения водоносные клетки передают воду главным образом ассимиляционным тканям. Мощно развитая водоносная ткань встречается у суккулентов ≈ растений с мясистыми сочными листьями и стеблями: агавы, алоэ, кактусы, молочаи. Развитой водоносной тканью снабжены клубневидные вздутия стеблей многих эпифитных орхидных. Однако подлинными рекордсменами в запасании влаги являются сфагновые мхи. Они могут накапливать такое количество влаги, которое в 40 - 50 раз превышает их сухой вес. Такая высокая гигроскопичность сфагнов связана с особенностями их анатомического строения. В листьях сфагновых мхов имеются две группы клеток: особые гиалиновые клетки, имеющие большие или многочисленные поры и накапливающие воду и обычные хлорофиллоносные клетки. Гиалиновые клетки занимают большую часть объема. Нередко крупные водоносные клетки имеют спиральные или кольчатые утолщения оболочек, придающих клеткам дополнительную прочность и предохраняющие их от слияния. Значительно шире распространены ткани, запасающие пластические вещества. Обычно запасы накапливаются в полостях клеток, реже в их оболочках. Наиболее распространенные запасные вещества: сахар, инулин, аминокислоты, белки, крахмал. В клеточных стенках обычно откладываются гемицеллюлозы. В этих случаях стенки чрезвычайно сильно утолщаются. В качестве примеров можно назвать запасающие ткани эндосперма семян кофе и финиковой пальмы. Воздухоносная паренхима в стебле рдеста блестящего (Potamogeton lucens) В растениях довольно часто встречается ткань с большим числом очень крупных межклетников — аэренхима (рис. 71). Соединяясь между собой, ее межклетники образуют единую вентиляционную сеть. Поэтому эту ткань иногда называют вентиляционной. Газовый состав межклетников значительно отличается от состава атмосферного воздуха, так как клетки в процессе своей жизнедеятельности выделяют в межклетники одни газы и поглощают другие. От условий обитания и особенностей строения того или иного растения зависит характер циркуляции газов по межклетникам, обеспечивающий нормальную жизнедеятельность растительного организма. Проходя по всему телу растения от корней до листьев, аэренхима выполняет вентиляционные и отчасти дыхательные функции, обеспечивая близлежащие ткани растения кислородом. Газы в аэренхиме перемещаются только путем диффузии. Аэренхиму можно рассматривать как модификацию паренхимы. Однако форма ее клеток очень разнообразна. Они могут быть округлыми (цветоножка кубышки), звездчатыми (стебель ситника) и др.  Рисунок 4. Воздухоносная паренхима в стебле рдеста блестящего (Potamogeton lucens) (по В.Х. Тутаюк, 1980): 1 — кутикула; 2 — эпидерма; 3 — клетки воздухоносной паренхимы; 4 — воздухоносная полость; 5 — эндодерма. Аэренхима — первичная ткань. Ее клетки имеют тонкие первичные стенки, покрытые со стороны межклетников слизью, уменьшающей транспирацию. Вакуоли крупные, цитоплазма занимает постенное положение и содержит много мелких лейкопластов. Аэренхима хорошо развита в органах водных и болотных растений: в цветоножках кубышки, стеблях белокрыльника, пушицы, рдеста, ситника, корнях камыша. У водных растений аэренхима выполняет не только вентиляционные функции. Воздушные полости в стеблях и листьях уменьшают удельный вес растений, позволяя им свободно плавать в воде. Водопоглощающая ткань (поглощающие, или всасывающие, ткани представлены ризодермой и веламеном). Ризодерма, или эпиблема, — первичная однослойная ткань. Она представляет собой наружный слой клеток молодых участков корней. Основная функция ризодермы — поглощение (всасывание) из почвенных растворов воды с растворенными в ней минеральными веществами. Через ризодерму из корня в почву могут выделяться некоторые вещества, например кислоты, которые, воздействуя на субстрат, делают его более пригодным для жизнедеятельности корней. Плотно сомкнутые клетки ризодермы тонкостенные, с вязкой цитоплазмой и большим числом митохондрий. Поглощающая поверхность ризодермы многократно увеличивается (в 10 раз и более) за счет образования корневых волосков. Корневой волосок представляет собой вырост клетки ризодермы длиной 1—3 мм (до 8 мм). При формировании корневого волоска наружная стенка клетки выпячивается и ядро перемещается в его растущую верхнюю часть. У одних растений все клетки ризодермы образуют корневые волоски, у других — уже на ранних этапах развития ризодермы выделяются клетки двух типов: способные образовывать корневые волоски трихобласты и не способные к этому атрихобласты. Веламен представляет собой многослойную ткань, покрывающую поверхность воздушных корней однодольных растений-эпифитов (орхидеи, бромелии). В результате быстрого отмирания протопластов плотно сомкнутые разные по форме клетки веламена заполнены воздухом, который во время дождей может замещаться водой. Вода поступает в клетки веламена не осмотическим путем (в них нет протопласта), а капиллярным — через крупные поры и перфорации в стенках наружных клеток. Стенки клеток веламена имеют сетчатые или спиральные утолщения, придающие им жесткость и предотвращающие их смыкание. Как поглощающие ткани можно рассматривать всасывающий слой клеток щитка (видоизмененной семядоли) зародыша злаков и внутренней поверхности ловчих аппаратов насекомоядных растений, а также гаустории (присоски) растений-паразитов. 40. Почки, строение и классификация. Биологическая роль почек. Почка - это зачаточный побег с очень укороченными междоузлиями (рис. 5). Центральную часть в почке занимает зачаточный стебель, на верхушке которого находится конус нарастания, представляющий собой образовательную ткань. На стебле расположены зачаточные листья. Снаружи почка покрыта почечными чешуями, способными предохранять зачаточные листья и конус нарастания от неблагоприятных условий среды. Для выполнения защитной функции у почечных чешуи образуется густое опушение, выделяются смолистые вещества и т. п. Весной почки деревьев и кустарников набухают, меристематические клетки конуса нарастания начинают интенсивно делиться, стебель трогается в рост, почечные чешуи раздвигаются и между ними появляются кончики зеленых листьев. Это происходит благодаря усиленному росту междоузлий. Одновременно разрастаются и листья, в результате чего формируется побег. Если побег развивается из почки в течение одного вегетационного периода, то его называют годичным побегом. 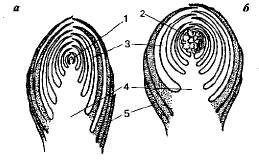 Рисунок 5. Схема строения вегетативной (а) и генеративной (б) почек (продольный разрез): 1 -- конус нарастания; 2 -- соцветие в зачаточном состоянии; 3 -- зачаточные листья; 4 -- зачаточный стебель; 5 -- почечная чешуя По внутреннему строению различают почки вегетативные и генеративные, или цветочные. В вегетативной почке расположены зачаточные стебель и листья. В цветочных почках кроме стебля и листьев имеются зачатки цветков (соцветий) или одного цветка; в последнем случае почку называют бутоном. Генеративные почки многих растений можно отличить от вегетативных не только по внутреннему строению, но и по внешнему виду: они более крупные, на верхушке округлые. По расположению на стебле различают верхушечные и боковые почки. Верхушечные почки находятся на верхушке побега и обеспечивают нарастание стебля в высоту (длину). Боковые почки закладываются в пазухах листьев (это пазушные почки) или вне пазухи на междоузлиях, корнях или листьях (придаточные почки) и развиваются в боковые побеги. В результате происходит ветвление побега и формируется верхняя часть растения, характерная для каждого вида. 58. Бесполое (спорами и зооспорами) и половое размножение (изогамия, гетерогамия, оогамия) растений. Привести примеры. Бесполое размножение — способ размножения, при котором потомство происходит от одного организма, а не от объединения гамет, как это осуществляется при половом размножении. Организмы, размножающиеся бесполым путем, могут страдать от опасного недостатка разнообразия — но они также могут размножаться быстрее, чем организмы, размножающиеся половым путем, и одна особь может основать новую популяцию без необходимости в спаривании. Некоторые организмы, практикующие бесполое размножение, могут обмениваться генетической информацией для повышения разнообразия, используя формы горизонтального переноса генов — например, бактерии, которые используют плазмиды для передачи небольших фрагментов ДНК. Однако этот метод приводит к появлению меньшего количества уникальных генотипов, чем половое размножение. Преимущества бесполого размножения К важным преимуществам бесполого размножения относятся: Быстрый рост популяции. Это особенно полезно для видов, чья стратегия выживания заключается в очень быстром размножении. Многие виды бактерий, например, могут полностью восстановить популяцию из одной мутантной особи в течение нескольких дней, если большинство членов популяции уничтожены вирусом. Для основания новой популяции не требуется спаривание. Это полезно для видов, члены которых могут оказаться в изоляции (например, грибов, растущих из спор, разносимых ветром; растений, чье половое размножение зависит от опылителей; животных, обитающих в среде с низкой плотностью популяции). Меньшие затраты ресурсов. Бесполое размножение, которое часто может осуществляться просто путем отщепления части родительского организма и обретения им самостоятельной жизни, требует меньше ресурсов, чем вынашивание нового детского организма. Например, многие растения и морские обитатели могут просто отрезать часть себя от родительского организма и заставить эту часть выжить самостоятельно. Таким способом можно получить только потомство, генетически идентичное родительскому организму: вынашивание нового организма, ткани которого отличаются от тканей родителей, требует больше времени, энергии и ресурсов. Эта способность просто разделиться на две части является одной из причин, почему бесполое размножение быстрее полового. Недостатки бесполого размножения Самый большой недостаток бесполого размножения состоит в отсутствии разнообразия. Поскольку члены популяции, размножающейся бесполым путем, генетически идентичны, за исключением редких мутантов, все они подвержены одним и тем же заболеваниям, дефициту питания и другим видам неблагоприятных факторов окружающей среды. Ирландский картофельный голод был одним из примеров отрицательной стороны бесполого размножения. Этот картофель, в основном, размножавшийся бесполым путем, оказался уязвимым, когда по острову прокатилась чума, убивающая картофель. В результате почти все посевы погибли, и многие люди умерли от голода. Другим примером является почти полное исчезновение банана Грос-Мишель — одного из двух основных сортов бананов в 20 веке — после появления болезни, к которой он был генетически уязвим, в результате чего стало невозможно выращивать его в коммерческих целях. С другой стороны, многие виды бактерий используют преимущества высокой частоты мутаций для создания определенного генетического разнообразия, используя бесполое размножение для быстрого роста своих колоний. У бактерий выше частота ошибок при копировании генетических последовательностей, что иногда приводит к созданию новых полезных признаков даже при отсутствии полового размножения. В природе встречаются примеры, когда особь способна размножаться двумя способами в зависимости от условий. Среди них — хламидомонада. Это одноклеточная зеленая водоросль. В благоприятных условиях она размножается бесполым путем благодаря зооспорам. Половое же размножение «включается» при неблагоприятных условиях: пересохший водоем, низкая температура снаружи и др. Типы бесполого размножения Существует множество различных способов бесполого размножения. Спорогенез — это производство репродуктивных клеток, называемых спорами, которые могут вырасти в новый организм. Споры часто используют стратегии, схожие со стратегиями семян. Но в отличие от семян, споры могут образовываться без оплодотворения половым партнером. Споры также чаще распространяются автономно, например, с помощью ветра, а не зависят от других организмов, например, животных-переносчиков. Споры, от «sporā», что означает «семя» и «genesis», что означает «рождение» или «происхождение», представляют собой спящие репродуктивные клетки, которые похожи на семена, служащие для рассеивания. Однако споры не являются семенами, так как в них отсутствует зародыш, образующийся при слиянии мужской и женской гамет. Споры имеют толстые стенки и очень устойчивы к различным неблагоприятным условиям, таким как высокая температура и низкая влажность. Когда условия подходят, они прорастают и дают начало новым особям. Сосудистые растения и грибы являются примерами бесполых организмов, которые размножаются путем образования спор. Зооспорангии — такой спороносящий орган, внутри которого образуются подвижные споры, снабженные жгутиками, — зооспоры. Зооспоры — это голые участки протоплазмы, лишенные оболочки, чаще с одним ядром, иногда с несколькими ядрами и с одним или двумя жгутиками. При помощи жгутиков зооспоры активно передвигаются в воде. Спорангий представляет собой спороносящий орган, в котором развиваются неподвижные споры, имеющие оболочку. Спорангиоспоры представляют собой многоядерные клетки, одетые оболочкой и лишенные органов движения. Эндогенное спороношение характерно для наиболее примитивных грибов. На нескольких примерах ознакомимся с особенностями бесполого размножения грибов. Гриб Olpidium brassicae вызывает заболевание рассады капусты, известное под названием черной ножки. Как осуществляется бесполое размножение у этого гриба? Его вегетативное тело в виде голого комочка протоплазмы располагается внутри клетки хозяина, т. е. гриб является эндобионтом. Это внутриклеточный паразит, который питается за счет содержимого клетки хозяина. Его протопласт постепенно разрастается, ядро многократно делится и он становится многоядерным. Вокруг этого комочка протоплазмы (амебоида) образуется оболочка. Содержимое внутри этой грибной клетки распадается на отдельные участки по числу ядер, снабженные одним жгутиком, при помощи которого осуществляется их передвижение. Вскоре в оболочке образуется выводной проток, который пробивает оболочку клетки-хозяина. Эти тельца через выводной проток выходят наружу в виде зооспор. Зооспора некоторое время плавает (при наличии капельножидкой влаги в почве), а затем прикрепляется к клетке корня хозяина, переливает в нее свое содержимое, и весь процесс начинается сначала. Здесь вся особь превращается в зооспорангий. Такие организмы, у которых все вегетативное тело превращается в зооспорангий, называются монокарпическими или холокарпическими. Synchytrium endobioticum — возбудитель рака картофеля, вызывает образование на клубнях опухолевидных вздутий. Как и Olpidium brassicae, он является внутриклеточным паразитом, вегетативное тело которого также представлено амебоидом — голым участком протоплазмы. В определенный период развития амебоид становится из одноядерного многоядерным, сильно разрастается, при достижении зрелости одевается внутри клетки-хозяина оболочкой и переходит в состояние покоя (стадия летней цисты), после которого изнутри его, прорывая оболочку, выступает пузырь, одетый собственной оболочкой, это — просорус. Внутри просоруса обособляются многоядерные участки протоплазмы, окруженные своими оболочками. Каждый такой многоядерный участок представляет собой зооспорангий. Образованная таким путем группа зооспорангиев получила название соруса зооспорангиев. В данном случае одна особь превращается в несколько зооспорангиев, и на их образование расходуется все вегетативное тело. У водных оомицетов зооспорангии развиваются на концах гиф мицелия. Примером может служить Saprolegnia. Его виды поселяются на трупах мелких животных, например насекомых, попавших в воду. Некоторые из них вызывают гибель мальков рыб. Белый мицелий, покрывающий этих животных, представлен разветвленными, неклеточными гифами, на концах которых и формируются зооспорангии. Их развитие происходит следующим образом. Отдельные ветви мицелия вздуваются, и вздутая часть отделяется от остальной грибницы поперечной перегородкой. В образовавшемся мешковидном отростке гифы происходит деление ядер, вокруг которых обособляется протоплазма и формируются двужгутиковые зооспоры. Зооспоры здесь двух типов: одни зооспоры грушевидные, они выходят из образовавшегося на вершине зооспорангия отверстия, некоторое время плавают, затем останавливаются, утрачивают жгутики и одеваются тонкой оболочкой. В таком состоянии зооспора находится в течение непродолжительного времени. Затем оболочка лопается и из нее выходит зооспора другой формы — бобовидной, жгутики которой прикреплены сбоку, один из них направлен вперед, другой — назад. Проплавав несколько минут, такая зооспора останавливается, округляется, одевается тонкой оболочкой, теряет жгутики и затем прорастает в мицелий. Такие дважды подвижные организмы (имеющие зооспоры двух типов) называют дипланетическими. Образование подобной формы является своеобразным приспособлением к их распространению, ибо при двойном плавании зооспора уйдет от материнского организма значительно дальше. Зооспорангии у видов Saprolegnia чаще всего одиночные, однако одно вегетативное тело здесь формирует не один зооспорангий, а несколько. Такие формы, у которых на одном вегетативном теле образуется много зооспорангиев, называют поликарпическими. Иногда у представителей Saprolegnia можно наблюдать интересное явление, когда происходит врастание нового зооспорангия в оболочку старого, опустевшего. Это явление получило название пролиферации зооспорангиев. В опустевший зооспорангии поступает, новая часть протоплазмы, которая одевается внутри него собственной оболочкой. Так может повторяться несколько раз. Подобная же картина наблюдается и у близких к пор. Saprolegniales представителей пор. Monoblepharidales, например у Monoblepharis regignens. В других случаях у Saprolegnia можно наблюдать образование на одной и той же ветви мицелия группы зооспорангиев в виде кустика. В этих случаях молодой зооспорангий закладыва ется непосредственно под уже имевшимся, вырастая под некоторым углом к последнему Эндогенными спорами являются также и спорангиоспоры формирующиеся внутри спорангия. Этот способ бесполого размножения наиболее распространен у мукоровых грибов (класс Zygomycetes). Спорангий развивается на спорангиеносце, представляющем собой вертикально расположенную гифу, отходящую от мицелия. В верхней части многоядерная клетка отде ляется поперечной перегородкой от остальной гифы. Эта клетка представляет собой будущий спорангий. В процессе роста она сильно вздувается, число ядер в ней увеличивается, затем вокруг нескольких ядер обособляется протоплазма, которая по периферии уплотняется, одевается оболочкой и образует неподвижные споры. При разрыве оболочки спорангия спорангиоспоры освобождаются и разносятся ветром. У многих мукоровых грибов верхняя часть спорангиеносца разрастается у углубляется внутрь спорангия, образуя колонку. Экзогенные споры (конидии) — это споры, образующиеся на поверхности спороносящего органа, на конидиеносцах, представляющих специализированные ветви мицелия. Конидии в простейшем случае развиваются на конидиеносце одиночно. При этом в верхней части конидиеносца отделяется спорогенная клетка, с увеличением своего размера она становится конидией. Затем перегородка между конидиеносцем и будущей конидией расслаивается, обе половины округляются, и конидия легко отпадает. На ее месте возникает другая конидия, и так происходит многократно. На одном конидиеносце в одних случаях развиваются одиночные конидии, в других случаях может образовываться группа конидий. Кроме того, конидии могут возникать и в цепочках. Образование цепочек может происходить двумя способами. В одних случаях это происходит так: верхняя клетка, отделившаяся от конидиеносца, растет и затем делится на две — верхнюю и нижнюю; вновь образовавшаяся верхняя клетка дает конидии, а нижняя, спорогенная, клетка вновь делится на две: верхнюю — конидию и нижнюю — спорогенную. Так наблюдается многократно, в результате образуется цепочка конидий, в которой самая верхняя конидия является наиболее зрелой. Такая цепочка распадается последовательно, начиная сверху (базипетальный тип). В других случаях происходит образование акропетальной цепочки, в которой наиболее старой является самая нижняя конидия, и распадается она только после созревания всех конидий. Из трех типов органов бесполого размножения, рассмотренных выше (зооспорангии, спорангии и конидии), наиболее примитивными следует считать зооспорангии, в которых формируются зооспоры, близкие по своему строению к тем простейшим формам, из которых возникают в последующем и растения, и животные. Спорангии и конидии несомненно являются более совершенными образованиями. Просматривая группу грибов в целом, мы можем обратить внимание на то, что все эти три типа органов бесполого размножения в своем происхождении тесно связаны друг с другом. На примере близких видов или одного и того же вида можно наблюдать случаи переходов от зооспорангия к спорангию и к конидии и от спорангия к конидии. Эволюция органов бесполого размножения у грибов теснейшим образом связана со средой обитания. При переходе грибов из водной среды обитания к условиям наземного существования у них появился ряд приспособлений, связанных с изменениями внешних условий среды. В первую очередь это отразилось на способах их размножения. Зооспорангии присутствуют преимущественно у водных грибов, однако они еще сохранились и у некоторых грибов, приспособившихся к наземному образу жизни, развивающихся на сухопутных растениях, в то время как спорангии и конидий характерны исключительно для наземных организмов. Большой интерес с точки зрения эволюции органов бесполого размножения представляет сем. Saprolegniaceae, объединяющее типичные водные организмы, развивающиеся на животных и растительных остатках, попавших в воду. Однако уже у этих форм намечается тенденция к сокращению подвижных стадий при формировании органов бесполого размножения. У типичного для этого семейства р. Saprolegnia, как уже отмечалось ранее, имеются дипланетические зооспоры, отличающиеся друг от друга по форме. Зооспоры первой фазы имеют грушевидную форму и два жгутика, которые располагаются у заднего конца. Такая зооспора, выйдя из зооспорангия, плавает 15—20 мин, иногда полчаса, затем утрачивая жгутики, облекается оболочкой, покоится некоторый промежуток времени, после чего прорастает. При этом происходит разрыв оболочки и изнутри выходит вновь зооспора, но уже бобовидной или почковидной формы и также с двумя жгутиками» но расположенными по-иному. Одни из них направлен назад, а второй — вперед. Таким образом, зооспоры первой и второй фаз резко отличаются друг от друга, хотя принадлежат к одному виду и развиваются у одного и того же организма. Такие зооспоры передвигаются активно и за двойное плавание уходят от материнского организма на значительное расстояние. Однако такие дипланетические формы встречаются не у всех представителей этого семейства. Так, у р. Achlya, например, первичные зооспоры, плавая 15—20 мин, могут затем одеваться оболочкой, после чего сразу прорастают в вегетативный росток грибницы. Вторая стадия развития и вторичные зооспоры здесь отсутствуют. Подвижность зооспор, следовательно, сокращена вдвое по сравнению с Saprolegnia. Представители другого рода этого семейства — Pythiopsis образуют также только один тип зооспор, т. е. здесь первичный процесс превращения зооспор первой фазы в неподвижные споры проходит внутри зооспорангия. Наружу выходят уже неподвижные споры, которые сейчас же в окружающей водной среде прорастают с образованием вторичных зооспор. Вторичные зооспоры вскоре прорастают в мицелиальный росток. У представителей Dictyuchus наблюдается еще более любопытное приспособление. Процесс превращения первичных грушевидных зооспор в неподвижные споры совершается и здесь внутри зооспорангия, который затем делится косыми перегородками на ряд ячеек, соответствующих количеству зооспор, развивающихся в таком зооспорангии. Для каждой зооспоры формируется свой собственный зооспорангий. А затем каждая из этих ячеек открывается своим отверстием, и оттуда выходят вторичные зооспоры, которые, находясь еще внутри зооспорангия, сбрасывают оболочки. В Traustotheca мы встречаемся с очень интересным явлением, когда из зооспорангия наружу высыпаются неподвижные споры, которые затем прорастают вегетативным ростком мицелия. Зооспорангий в этом случае по своему внешнему облику и по своему содержимому превратился в спорангий, внутри которого развиваются неподвижные споры. Разница между зооспорангием и типичным спорангием заключается в том, что споры в типичном спорангии имеют свою собственную оболочку. Здесь же, у Traustotheca, эта оболочка появляется после нападения внутреннего содержимого зооспорангия на первичные зооспоры. Из этого примера видно, что совершенно несомненна связь настоящих зооспорангиев, заполненных (в случае Traustotheca) неподвижными спорами, с типичными спорангиями, внутри которых с самого начала возникают неподвижные споры. Наконец, в пределах этого же семейства, у видов р. Aplanes. зооспоры претерпевают все фазы развития вплоть до образования вегетативного ростка внутри зооспорангия. Связь таких органов спороношения, как зооспорангий и конидии, можно проиллюстрировать на примере представителя класса Oomycetes — Phythophthora infestans, возбудителя фитофтороза картофеля. Этот гриб является типичным наземным организмом, паразитирующим на различных органах растений из сем. пасленовых — картофеле, томатах и др. Бесполое размножение этого организма осуществляется при помощи зооспорангиев, которые часто называют конидиями. И то и другое название к этим органам вполне применимо, так как один и тот же орган может функционировать либо как зооспорангий, либо как конидия. Phytophthora infestans имеет эндофитную грибницу. Спороносящие органы — спорангиеносцы, или конидиеносцы, — образующиеся на эндофитной грибнице, выходят наружу через устьица с нижней стороны пораженного листа. На кончиках спорангиеносцев развиваются лимонообразные зооспорангии или конидии. Если поместить такие образования во влажную камеру на предметное стекло в каплю воды и наблюдать за их поведением, то через несколько часов мы увидим, что на кончике зооспорангия (или конидии) произойдет разрыв оболочки с образованием отверстия и изнутри наружу выйдут типичные бобовидные зооспоры с двумя жгутиками. В каждом зооспорангии формируется от 8 до 16 зооспор, которые после выхода из зооспорангия активно плавают в капле воды. Это типичный зооспорангий. Проведем с тем же самым зооспорангием следующий эксперимент. Поместим зооспорангий на предметное сухое стекло, но создадим некоторую влажность в окружающей среде (покроем стеклянным колпаком со смоченной водой фильтровальной бумагой). Вместо зооспор из такого зооспорангия вырастет ростковая трубка, которая затем удлинится в вегетативный росток. Это типичная конидия. В разных условиях одно и то же образование — зооспорангий — ведет себя по-разному: либо как типичный зооспорангий, либо как конидия. Точно так же ведут себя зооспорангии целого ряда других родственных организмов в пределах близких семейств пор. Peronosporales. Представители сем. Pythiaceae — в основном сапрофитные формы, некоторые из них являются водными обитателями. Среди литиевых есть и фитопатогенные организмы, вызывающие болезни растений. Бесполое размножение у них осуществляется с помощью зооспор, развивающихся в зооспорангиях, которые образуются на концах вегетативных гиф, т. е. на недифференцированных ветвях мицелия. У одних видов зооспорангий прорастает, находясь еще на мицелии, у других видов зооспорангии вначале опадают, и только после этого начинается их прорастание. Это первый шаг на пути перехода к конидиальному спороношению. При прорастании такой зооспорангий дает трубку, заканчивающуюся пузырем, и уже в этом пузыревидном вздутии происходит формирование зооспор. У более высокоорганизованных представителей пор. Peronosporales мы отмечаем дальнейшие этапы на пути замены зооспорангиев конидиями. Во-первых, у них закрепляется свойство зооспорангиев отпадать после созревания и переноситься ветром на другие субстраты и только после отпадения от спорангиеносцев переходить к прорастанию (виды р. Phytophthora, виды р. Plasmopara). Во-вторых, образование зооспорангиев начинает происходить у многих форм не на ветвях мицелия, а на специализированных частях грибницы, т. е. на спорангиеносцах, которые у более примитивных форм хотя морфологически и отличаются от вегетативной грибницы, однако функционально еще не отделены от нее, так как сохранили способность к продолжению роста даже после формирования и опадения зооспорангиев (например, Phyiophtliora). Наконец, у наиболее высокоорганизованных представителей пор. Peronosporales (Plasmopara, Peronospora, Bremia) наблюдается как морфологическая, так и функциональная дифференциация спорангиеносцев (Plasmopara) и полная замена их конидиеносцами, формирующими типичные конидии (Bremia, Peronospora). Конидиеносцы здесь по форме и строению четко отличаются от мицелия, являясь специализированными органами, так как прекращают свой рост и развитие после образования конидий. У большинства грибов сем. Peronosporaceae, как правило, имеются типичные зооспорангии. Но у наиболее высокоорганизованных форм, таких, как Bremia и Peronospora, подвижные стадии в развитии органов бесполого размножения полностью утрачены. У этих форм зооспорангиев совершенно не образуется, хотя по развитию органов спороношения и другим признакам они близки к тем формам, которые имеют зооспорангии. Очень любопытно, что у конидий Bremia мы наблюдаем еще наличие сосочка, через который происходит их прорастание, тогда как у видов Peronospora прорастание совершается в любом месте конидии. Это наиболее высоко эволюционировавшие формы, приспособившиеся исключительно к паразитированию на наземных растениях. На этом основании мы можем считать, что зооспорангий является родоначальником конидии, а конидии, по крайней мере впервые возникшие, происходят от зооспорангиев, получивших при определенных условиях способность прорастать вегетативным ростком. У представителей сем. Albuginaceae (паразиты высших растений) пор. Peronosporales бесполое размножение осуществляется с помощью зооспорангиев, развивающихся на коротких спорангиеносцах. Они располагаются на мицелии на ограниченном участке субстрата в виде ложа из тесно примыкающих друг к другу спорангиеносцев. Ложе развивается вначале под покровом ткани растения-хозяина и обнажается только при полном его созревании. Это первый случай возникновения ложа у грибов. Видимо, оно оказалось биологически целесообразным, так как затем находит свое развитие у многих более высокоорганизованных грибов. Рассмотрим переход спорангиев к структурам, функционирующим как конидии. Спорангии, как известно, характерны исключительно для мукоровых грибов (пор. Mucorales), у которых на ценотическом, неклеточном мицелии образуются спорангиеносцы со спорангием на вершине, внутри которого формируется большое количество спорангиоспор. Созревшие в спорангиях спорангиоспоры при разрушении верхних частей оболочек спорангия высыпаются и разносятся ветром. У видов наиболее широко распространенного р. Mucor формируются только такие многоспоровые спорангии. У Thamnidium мы наблюдаем несколько иную картину. Здесь, так же как и у Mucor, возникает многоспоровый спорангий, располагающийся на вершине спорангиеносцев в виде вздутой клетки, внутри которой развиваются в большом количестве спорангиоспоры. Однако кроме многоспорового спорангия на боковых ответвлениях спорангиеносцев развиваются еще и малоспоровые спорангии — спорангиоли. Спорангиолей всегда развивается много, но количество спор в них не более 2—6. У близкого к Thamnidium другого рода мукоровых грибов — Chaetocladium, паразитирующего на других мукоровых, многоспоровый спорангий совсем не формируется. На боковых ответвлениях спорангиеносца развиваются только малоспоровые спорангии — спорангиоли, внутри которых, как правило, образуется лишь по одной спорангноспоре, при этом се оболочка часто срастается с оболочкой спорангиоли, так что по существу это образование можно рассматривать как конидию. Спорангиоли и ведут себя, как конидии: могут отрываться от конца спорангиеносца, разноситься воздушными течениями и прорастать вегетативным ростком грибницы. Таким образом, путем уменьшения спор внутри споровместилища, вплоть до одной, и происходит превращение спорангиев в конидии. Переход от спорангиев к конидиям у мукоровых грибов может осуществляться и другими путями. Так, например, у представителей Cunninghamella на начальной стадии имеет место образование типичного шаровидного вздутия на вершине спорангиеносца. Но развитие спорангия на этом и закапчивается, внутри него спор не возникает. Затем на оболочке спорангия образуются небольшие выросты, которые называются стеригмами, и на их концах затем и развиваются споры. Таким образом, в этом случае спорообразование вынесено изнутри на поверхность того же спорангия, хотя споры приобрели новый характер и оказались не эндогенными, каким являются спорангиоспоры, а экзогенными, т. е. конидиями. У представителей Syncephalis и Piplocephalis на расширенной части спорангиеносца образуются цилиндрические спорангиоли, и освобождение спорангиоспор здесь происходит либо после разрушения оболочки спорангиолей, либо после распадения этих спорангиолей на отдельные клетки. Таким образом, несмотря на принципиальное различие в типах бесполого размножения грибов (зооспорангий, спорангий, конидия), между ними существует тесная связь их друг с другом, и это дает нам возможность понять ход эволюционного процесса в прошлом. То, что зооспорангии в процессе эволюции могли быть заменены спорангиями, легко объясняется влиянием окружающей среды, связанным с переходом водных организмов к наземным, более сложным условиям существования. Прежде чем перейти в эти новые условия и приобрести новые свойства, организмы должны были иметь и потенциальную возможность приспособления к этим условиям. В превращении зооспорангия в спорангий мы видим только повышение качества спороносящих органов. Спорангии являются более совершенными, они лучше приспособлены к наземным условиям, чем зооспорангии: спорангиоспоры, снабженные оболочкой, хорошо защищены от высыхания и легко разносятся потоками воздуха. Повышение качества при превращении зооспорангия в спорангий произошло без утраты каких-либо свойств, поскольку и внутри зооспорангия, и внутри спорангия образуется большое количество спор. Такое превращение несомненно оказалось целесообразным и привело к формированию и расселению наземных организмов. Но когда происходит замена типичного зооспорангия конидией или спорангия конидией, то здесь наряду с приобретением нового качества при переходе из водной среды в сухопутную одновременно наблюдается как бы редукционный процесс в виде уменьшения количества спор. Так, у Phytophthora infestans внутри типичного зоспорангия развивается до 10—16 зооспор, из которых каждая может дать начало новому организму. А когда у этого же вида зооспорангий функционирует как конидия, то возникает только один вегетативный росток при ее прорастании, который дает начало лишь одной особи. 1 2 |