бактериотоксины. Бактериальные токсины
 Скачать 143.5 Kb. Скачать 143.5 Kb.
|

|
БАКТЕРИАЛЬНЫЕ ТОКСИНЫ Классификация бактериальных токсинов. Луи Пастером в 1887 г. были проведены опыты доказывающие, что веществами, которые образуются в результате жизнедеятельности микроорганизмов и находятся в питательном бульоне, можно вызывать такие же клинические признаки заболевания, как и при заражении самим возбудителем болезни. Э. Ру и А. Иерсин в своих экспериментах 1889 года подтвердили этот вывод. Дальнейшие исследования показали, что безмикробные, стерильные фильтраты, полученные с жидких питательных сред, где размножались изучаемые микроорганизмы, вызывают клинические проявления и патологические изменения, характерные для столбняка, ботулизма, холеры, скарлатины. Познее выяснилось, что проявление ряда патологических процессов при многих инфекционных заболеваниях вызвано продуктами жизнедеятельности микробов. Эти продуцируемые микроорганизмами вещества получили название микробных токсинов. Уже к 1890 году были обнаружены токсины двух важнейших патогенных для человека микроорганизмов (вызывающих большой процент смертельного исхода при заболевании) С.diphtheriae - дифтерия и Cl.tetani - столбняк. Постепенно, в ходе экспериментов, все токсины микробной этиологии разделили на две группы. В первую включили токсические продукты связанные со стромой (телом) микробной клетки. Они становятся токсичными только после гибели и разрушения микроорганизмов. Эту группу токсинов выявили у грамотрицательных бактерий и назвали эндотоксинами (эндо - endo - внутри). Для них характерна низкая специфичность действия. При введении экспериментальным животным, все они вызывают схожие клинические и патологические симптомы. Изучение их природы и места локализации в клетке потребовало длительного, интенсивного исследования. Сейчас установлено, что эндотоксины – это комплекс липополисахаридов с белками, которые находятся в наружных слоях клеточной стенки грамотрицательных бактерий. Во вторую группу отнесли секретируемые или растворимые микробные токсины. Они выделяются в окружающую среду при жизни микроорганизмов и не связаны со стромой последних. Эти токсины оказались чувствительны к нагреванию и являются белками. Так как они присутствуют в среде и не являются структурной частью микроорганизма, то получили название экзотоксины (экзо – exo – снаружи, вне). В экспериментах было доказано, что экзотоксины оказывают специфическое действие на организм, характерное для той или иной болезни. Термины «эндотоксины» и «экзотоксины», которыми называют две выше указанные группы токсических веществ не должны вводить в заблуждение. В настоящее время есть данные, показывающие, что многие «экзотоксины» связаны с бактериальными клетками во время их роста и высвобождаются только после гибели и лизиса (разрушения) бактерий. Общепринято что, экзотоксины являются белками, а эндотоксины - молекулярными комплексами, содержащими белок, липид и полисахарид. Приведенные выше термины, в настоящее время настолько общеприняты, что отказываться от них никто не хочет. Предложенный М. Далиным и Н. Фишем (1980) термин «мезотоксины», объединяющий те токсины микроорганизмов, что имеют общие характеристики первой и второй групп, не получил признания. В таблице 1 дана дифференциальная характеристика экзо- и эндотоксинам. Таблица 1 Сравнительная характеристика экзо- и эндотоксинов. (по Н. Колычеву, 1991г.)  . . Однако, биохимики, разделение микробных токсинов на группы, проводят в соответствии с данными об их природе и химических свойствах. Они различают группу простых и сложных белков (протеотоксины), группу со стероидной конфигурацией (афлотоксины) и группу липополисахаридных комплексов, токсическую активность которых определяет липидный компонент (липид А). Такой биохимический, а в последствии иммунохимический подход позволил теоретически и биохимически обосновать получение антитоксинов (Э. Беринг, 1892г.). С их помощью смогли отличать один токсический микробный биополимер от другого, микробиологи – отличать in vitro (в пробирке) токсигенные штаммы от нетоксигенных, патофизиологи – понимать (в какой-то степени) механизм поражающего действия токсинов. П. Эрлих (P. Ehrlich), используя антитоксины, как молекулярные зонды, впервые выявил молекулярную характеристику микробных токсинов. Позднее Г. Рамон (G. Ramon) опираясь на его теоретические разработки, организовал производство анатоксинов. Исследования по антитоксинам позволили провести разделение, дифференциацию токсинов на серотипы (серогруппы, сероварианты) в соответствии с их антигенной структурой. Однако при серологическом анализе доказана некоторая идентичность токсинов, вырабатываемых бактериями разных видов и родов. Выяснилось, что антигенно-родственными оказались холерный токсин и термолабильные энтеротоксины, продуцируемые E.coli, Sal.typhimurium. Установлена антигенная похожесть энтеротоксинов выделяемых бактериями видов Sh.plexneri и Sh.dysenteriaе. Высокая степень сходства отмечается у токсинов (гемолизина) Cl.tetani, Bac.cereus и Diplococcus pneumonie и St.pyogenes, СL.perfringens. Проведенные иммунологические исследования позволяют рассматривать микробные токсины как совокупность серогрупп, которые сходны в каких-то структурах по своему молекулярному строению. Стало ясно, что микробные токсины можно группировать не только по сходству происхождения (эндо- или экзо-), химической природе (белки, липополисахариды), по сходству в молекулярной организации, по его антигенной структуре, но и потому, какую роль играют отдельные структурные единицы или молекулы в патогенезе интоксикации на клеточном или субклеточном уровнях. Так, например выявлено, что экзотоксины шигелл, синегнойной палочки, дифтерийной бактерии почти одинаковым образом блокируют синтез белков на субклеточном уровне, причем два последних выводят из строя один и тот же фермент - трансферазу II. Обнаружилась функциональная общность холерного токсина и термолабильного токсина E.coli, поражающее действие которых связано со способностью активировать клеточную аденилатциклазу. Благодаря методическому подходу по разделению (дифференциации) микробных токсинов по указанному принципу были уточнены некоторые особенности микробных токсинов. Получены данные, что они отдельными участками своих молекул иммитируют (подражают) структуре ферментов, гормонов, нейромедиаторов макроорганизма. Возможно, эта особенность и обеспечивает микробным токсинам способность вмешиваться в обменные процессы у макроорганизма (человека). Патогенез (болезнетворность), бактериальных токсинов. Опираясь, на наиболее распространенную, схему дифференциации бактериальных токсинов рассмотрим, ту роль, которую они играют в патогенезе как при токсикоинфекциях и токсикозах человека, так и при других инфекционных болезнях. Патогенетическое действие экзотоксинов достаточно наглядно представлено в таблице 2 (R.Stanier et.all,1976г.) Таблица 2 Основные экзотоксины и их патогенетическое воздействие. (по R. Stanier. et. all, 1976 г.) 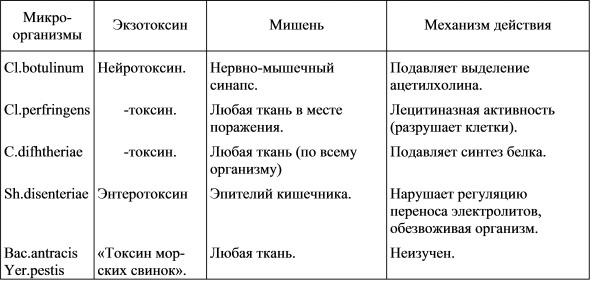 Те токсины, которые выделяются во время роста грамположительными бактериями, являются истинными экзотоксинами. Их не обнаружили не в цитоплазме, не в структуре тела. По всей вероятности их синтез происходит на связанных с мембраной рибосомах. Однако механизм, с помощью которого эти белки выходят сквозь клеточную стенку наружу, полностью неясен. Неясна до конца и функция экзотоксина. Установлено, что многие не токсигенные штаммы размножаются в клетке хозяина так же интенсивно, как и токсигенные. Причем гены, определяющие синтез экзотоксина, часто локализованы в плазмидах или профагах, а не на бактериальной хромосоме. Сейчас выяснено, что дифтерийный токсин, токсин Str.pyoqenes, энтеротоксин Staph. aureus, токсин Cl.botulinum типа «D» детермируется генами профага, а некоторые токсины E.coli - плазмидными генами. Потеря данного генетического материала ведет к утрате способности продуцировать токсин. При введении в бактериальную клетку такого детерменированного профага или плазмиды, образование токсина восстанавливается. Другим примером иллюстрирующим выше проведенный тезис, могут служить токсины Cl.botulinum типа «А» и «В», а также Е- и i-токсины Cl.perfringens. Они образуются в результате протеолитического расщепления более крупных нетоксичных полипептидов. Для этого необходимо, чтобы протеолитические ферменты, расщепляющие их, выделялись бактериями в культуральную среду (жидкая питательная среда для размножения бактерий). МЕХАНИЗМ ДЕЙСТВИЯ ЭКЗОТОКСИНОВ проявляется вмешательством в работу определенных функций, тканевых клеток или в разрушении некоторых субклеточных структур. Примером первого может служить дифтеритический токсин, продуцируемый Cor.diphthеriae. Он влияет на процессы синтеза ДНК, РНК и белка клетки. Подавление этих процессов происходит в результате того, что токсин инактивирует фермент трансферазу II. А так как этот фермент способствует переносу растущей полипептидной цепи с одной молекулы т-РНК на другую на поверхности рибосом, то разрушение трансферазы II и останавливает синтез белка. Примером разрушения субклеточных структур бактериальным токсином может служить токсин Cl.perfringens. По своей природе это лецитиназа. Она расщепляет лецитин, являющийся важным структурным компонентом клеточной мембраны. Его расщепление приводит к разрушению мембраны разнообразных тканевых клеток, что возможно и является причиной распада тканей при газовой гангрене. Лецитиназу продуцируют и микроорганизмы родов Bacillus, Staphyloccocus, входящие в группу пищевых токсикозов. Представители группы пищевых токсикоинфекций (например: E.coli, Sal.typhimurium, Cl.perfringens), а так же возбудители холеры и бактериальной дизентерии продуцируют энтеротоксины (entero - кишка), которые специфически действуют на эпителий кишечника. Механизм их действия заключается в том, что эти вещества связываются со специфическими рецепторами мембраны эпителия. Связанный токсин активирует мембранную аденилатциклазу, это вызывает повышение концентрации в клетке циклического аденозинмонофосфата (АМФ), что в свою очередь вызывает повышение скорости переноса электролитов, т.е. утечку из тканевых структур. Вместе с ними уходит и вода. В результате происходит потеря тканевой жидкости, а это приводит к обезвоживанию организма и шоку. Если не восполнить потерю жидкости и электролитов циркулирующих в организме, то наступает смерть. Механизм патогенетического действия эндотоксинов иной. Как уже указывалось выше, по своей химической природе это комплекс липополисахаридов с белками клеточных стенок грамотрицательных бактерий. Таким образом они идентичны О -антигенам (соматическим антигеном) целой клетки. Эндотоксины выделены из всех патогенных граммотрицательных бактерий. Для токсинов этой группы характерны два типа механизма активности - они вызывают повышение температуры тела (пирогенность) и являются токсичными. Этими двумя свойствами обладает липополисахаридная фракция токсина, белковая фракция имеет только антигенные свойства. Пирогенная активность липосахаридного комплекса высока. Введение его лошади весом в 700 кг в количестве 0,000001 г вызывает повышение температуры тела. Известно , что температура животного, человека регулируется определенными центрами в головном мозге. Однако , эндотоксины не действуют на эти центры. Они действуют на полиморфноядерные лейкоциты, вызывая освобождение пирогенного вещества. Химическая природа его до настоящего времени точно не ясна, но установлено, что оно и вызывает повышение температуры. Патогенетическое действие данной группы токсинов заключается и в том, что они увеличивают проницаемость капилляров и вызывают разрушение клеток, в свою очередь выделяются воспалительные агенты, которые так же участвуют в развитие патологического процесса. Механизм воспалительного процесса до конца не известен, возможно тут значительную роль играет не токсин (липополисахаридная часть эндотоксина), а аллерген - т.е. антигенный продукт микроорганизма (соматическая часть клетки), который индуцирует воспалительную аллергическую реакцию у сенсибилизированного макроорганизма. |