Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.1 58 ГЛАВА 5
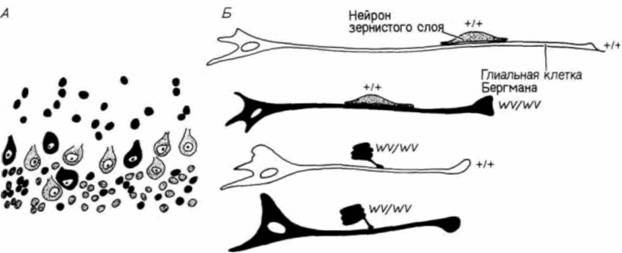
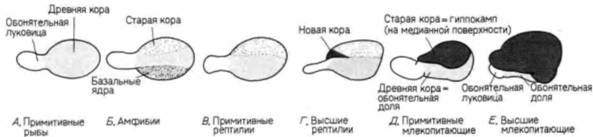
ден с фенотипическим проявлением мутации staggerer; строение клеток Пуркинье у них также аномально. Однако у мышей weaver остановка роста клеток Пуркинье представляет собой вторичное явление — она обусловлена, по-видимому, генетическим дефектом клеток зернистого слоя. Это было доказано в опытах «конструирования» химерных мышей из зародышей дикого типа и мутантов weaver (Goldowitz, Mullen, 1982). У химерных мышей клетки дикого типа можно обнаружить в мозжечке по низкому содержанию в них ß-глюкуронидазы (выявляемой гистохимическим методом) и по иной форме ядер. У мутантов меньше клеток Пуркинье, чем у мышей дикого типа, и локализация этих клеток в коре мозжечка аномальна. У химерных зародышей некоторые из нейронов Пуркинье также располагались не там, где они должны были находиться. Эти эктопические нейроны представляли собой клетки как того, так и другого генотипа. Однако все клетки зернистого слоя у мышей дикого типа находились там, где им положено, тогда как у мутантных зародышей эти клетки были случайно распределены по коре мозжечка (рис. 5.17. А). Таким образом, дефект у мышей weaver заключается, по-видимому, в отсутствии у клеток зернистого слоя способности мигрировать из наружного герминативного слоя в ту область коры мозжечка, где они должны находиться. В зернистый слой нейробласты обычно мигрируют, двигаясь по длинным отросткам глиальных клеток Бергмана (Rakic, Sidman, 1973). Хаттеном и его коллегами (Hatten et al.. 1986) было показано, что у мутантов weaver нейробласты неспособны узнавать клетки Бергмана как субстрат, по которому они должны мигрировать. В опытах этих исследователей в разных сочетаниях соединяли клетки зернистого слоя и клетки глии нормальных и мутантных зародышей (рис. 5.17, Б). Клетки от мышей дикого типа прочно слипались с глией зародышей обоих генотипов (как это обычно и бывает, когда клетки мигрируют). Однако у мутантных зародышей они не приближались ни к глиальным клеткам зародышей дикого типа, ни к глиальным клеткам зародышей weaver. Эти данные убедительно подтверждают предположение, что генетический эффект мутации weaverзаключается в неспособности будущих клеток зернистого слоя узнавать клетки Бергмана и мигрировать по их поверхности. Трехзонная структура мозга в полушариях также претерпевает существенные изменения. Некоторые нейробласты из плащевого слоя мигрируют через белое вещество и образуют вторую зону нейронов, называемую новой корой (неопаллиумом, или неокортексом). Эта новая кора мозга затем подразделяется на шесть слоев, содержащих тела нейронов. Дифференцировка этих слоев продолжается после рождения ребенка в течение первой половины детства. Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.РАННЕЕ РАЗВИТИЕ ПОЗВОНОЧНЫХ. ЭКТОДЕРМА 159 Дополнительные сведения и гипотезы: Эволюция развития головного мозгаЭволюция коры головного мозга — один из наиболее интересных разделов анатомии позвоночных. Доли мозга возникают как парные выросты переднего мозга. У рыб, например, они первоначально функционируют как органы обоняния. У амфибий и рептилий древнюю кору (палеопаллиум) головного мозга покрывают старая кора (архипаллиум) и полосатое тело (corpus striatum) (рис. 5.18). Архипаллиум. по-видимому, связан с «эмоциональными» типами поведения, тогда как палеопаллиум и полосатое тело имеют отношение к автоматическим «инстинктивным» реакциям. У птиц полосатое тело развито очень хорошо. У более высокоорганизованных рептилий появляются первые нейроны, связанные с новой корой (неопаллиумом, или неокортексом). Эволюция мозга млекопитающих, и особенно человека, характеризуется необычайно интенсивным ростом и распространением новой коры, оттесняющей внутрь архипаллиум, который формирует гиппокамп (ответственный за сексуальное и агрессивное поведение). Мозолистое тело функционирует как реле для переключения некоторых непроизвольных реакций. Неокортекс становится центром обучения, памяти и интеллекта. У птиц нет новой коры, и у видов, способных к обучению, его функцию выполняет гиперстриатум (Romer, 1976). Развитие новой коры у человека занимает чрезвычайно большой период времени. Фактически у ребенка после его рождения мозг продолжает развиваться с такой же скоростью, как у плода (Holt et al., 1975). Портманн (Porlmann. 1941. 1945) полагает, что по сравнению с другими приматами у человека очень короткий период беременности. Сравнивая развитие плода у человека и других приматов, он определил, что беременность у женщины должна была бы продолжаться 21 мес. Портманн и Гоулд (Gould. 1977) считали, что именно такова и есть продолжительность беременности у человека, поскольку все мы в течение первого года жизни являемся в сущности внематочными плодами. Это утверждение основывается на том, что и после рождения мозг ребенка продолжает расти чудовищно быстрым темпом, характерным для плода. Ни одна женщина не могла бы родить 21-месячного младенца, потому что его голова не могла бы пройти через родовые пути. Роды должны состояться в то время, когда голова у младенца еще достаточно мала, чтобы пройти через материнский таз. Поэтому человеческие детеныши рождаются в конце девятого, а не двадцать первого месяца беременности. Кроме того, именно в течение этого времени мы впервые подвергаемся влиянию окружающего мира. Наша развивающаяся нервная система получает существенный выигрыш от воздействия на нее столь сильной стимуляции в течение первого года жизни ребенка.
|