Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.20________________ ГЛАВА 1________________________________________________________________________________
Происхождение полового размноженияПоловое размножение это еще одно изобретение простейших, которое оказало глубокое воздействие на более сложные организмы. Необходимо отметить, что половой процесс и размножение - суть два различных явления, которые могут существовать отдельно друг от друга. Размножение - это возникновение новых особей, половой процесс - это создание новых комбинаций генов, происходящих οт двух разных особей. Размножение при отсутствии полового процесса свойственно тем организмам, которые размножаются простым делением: когда происходит деление амебы или отпочковывание новых особей гидры, никакого перераспределения генов не наблюдается. У одноклеточных организмов нередко
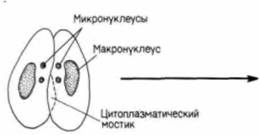
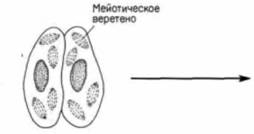
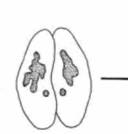
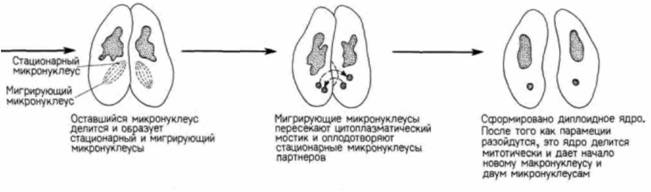
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.ВВЕДЕНИЕ В ИЗУЧЕНИЕ РАЗВИТИЯ ЖИВОТНЫХ____________________________________________ 21 встречается также половой процесс без размножения. Бактерии могут передавать гены от одной особи другой с помощью особых половых ворсинок, называемых половыми пилями или фимбриями (рис. 1.12). Эта передача осуществляется независимо от размножения. Простейшие также способны перераспределять гены независимо от размножения. Так, например, у парамеций размножение происходит путем простого деления надвое, а половой процесс путем конъюгации (рис 1.13). Две парамеции соединяются своими ротовыми аппаратами, и между ними возникает цитоплазматический мостик. У обеих парамеций макронуклеус (осуществляющий регуляцию метаболических процессов) разрушается, тогда как микронуклеус подвергается мейозу, за которым следует митоз; возникает восемь гаплоидных микронуклеусов, из которых все, за исключением одного, разрушаются. Оставшийся микронуклеус делится снова и образует два микронуклеуса стационарный и мигрирующий. Каждый мигрирующий микронуклеус перемещается по цитоплазматическому мостику в соседний конъюгат и сливается с его стационарным микронуклеусом («оплодотворяет» его), благодаря чему в обеих клетках возникает новое диплоидное ядро. Когда конъюгирующие партнеры расходятся, это диплоидное ядро делится, давая начало новому микронуклеусу и новому макронуклеусу. При этом парамеции не размножаются, происходит только половой процесс. У одноклеточных эукариот наблюдается и объединение этих двух независимых явлений, полового процесса и размножения; в этом случае говорят о половом размножении. На рис. 1.14 представлен жизненный цикл хламидомонады (Chlamydomonas). Этот организм обычно существует в виде гаплоидной формы подобно гаметам млекопитающих, т.е. каждая хромосома у хламидомонады имеется в единственном числе. Однако особи каждого вида по поведению при спаривании подразделяются на две группы - плюс и минус. Когда особи разных групп встречаются, происходит объединение их цитоплазмы и слияние ядер с образованием диплоидной зиготы. Эта зигота является единственной диплоидной клеткой в жизненном цикле хламидомонады: осуществляя в конечном счете мейоз, она образует четыре новые клетки. Здесь мы имеем дело с половым размножением, поскольку в процессе мейоза происходит перераспределение хромосом, и вместе с тем возникает большее число особей. Обратите внимание, что при таком типе полового размножения простейших гаметы морфологически идентичны - различия между спермием и яйцом еще не возникло. При возникновении в эволюции полового размножения прогресс был достигнут в двух отношениях. Во-первых, возник механизм мейоза (рис. 1.15). с помощью которого диплоидный набор хромосом редуцируется до гаплоидного состояния (этот процесс будет подробно обсуждаться в гл. 22). Во-вторых, возник механизм, благодаря которому особи двух типов, различающиеся по полу, узнают друг друга. Узнавание происходит первоначально на уровне мембран жгутиков (рис. 1.16.; Goodenough. Weiss. 1975; Bergman et al.. 1975). Агглютинация жгутиков делает возможным установление контакта между особыми участками на клеточных мембранах Эти специализированные участки содержат специфические для особей разного типа компоненты, благодаря которым осуществляется слияние цитоплазмы этих особей. После агглютинации жгутиков плюс-особи инициируют слияние, образуя «трубку оплодотворения», сходную с той, которую мы встретим у спермиев. Эта трубка контактирует и сливается со специализированным участком на поверхности минус-особи. Интересно, что тот же механизм, который используется для выдвижения этой трубки,
|