Гилберт С. Биология развития. Т.1.doc , БИР. Библиография Гилберт С. Биология развития в 3х т. Т. I Пер с англ. М. Мир, 1993. 228 с
 Скачать 10.9 Mb. Скачать 10.9 Mb.
|

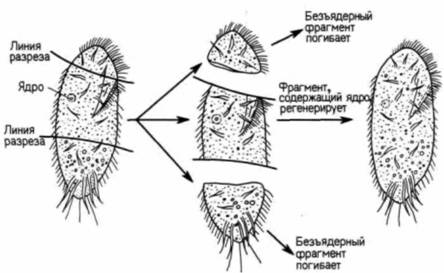
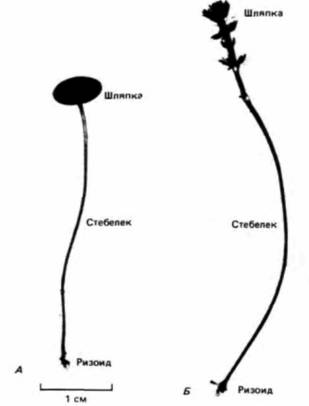
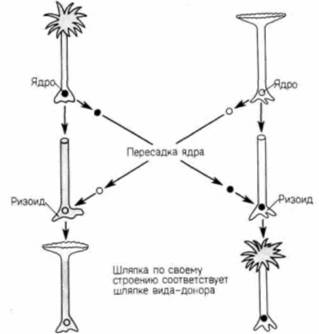
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.16_______________ ГЛАВА 1________________________________________________________________________________ полимеразами. У прокариот новосинтезированная мРНК немедленно начинает транслироваться с образованием белка, в то время как другой конец молекулы еще продолжает транскрибироваться с ДНК (рис. 1.4). Таким образом, транскрипция и трансляция у прокариот осуществляются одновременно и координированно. Существование ядерной оболочки у эукариот делает возможным совершенно новый тип клеточной регуляции. Рибосомы, ответственные за трансляцию, находятся по одну сторону ядерной оболочки, тогда как ДНК, а также РНК-полимераза, необходимые для трансляции, по другую ее сторону. В период между транскрипцией и трансляцией транскрибированная РНК должна претерпеть такие изменения, которые позволили бы ей пройти через ядерную оболочку. Регулируя выход мРНК в цитоплазму, клетка способна отобрать те из вновь синтезированных носителей информации, которые должны транслироваться. Таким образом, сложность организации достигает нового уровня, который, как мы увидим, чрезвычайно важен для развивающегося организма. Развитие одноклеточных эукариотВсе многоклеточные эукариотические организмы произошли в процессе эволюции от одноклеточных простейших. У этих простейших впервые появились основные особенности, присущие развитию эукариот. Примитивные эукариоты дают первые примеры морфогенеза, контролируемого ядром, использования клеточной поверхности как посредника для кооперации отдельных клеток, появления полового размножения. Регуляция морфогенеза при развитии ацетабулярииВ конце XIX в. еще не было доказано, что в ядре содержится информация, определяющая наследственность и направляющая развитие. Одним из лучших свидетельств в пользу такого представления были результаты опытов по разделению одноклеточных организмов на фрагменты, содержащие ядро, и безъядерные (Wilson. 1896). Когда различных простейших разрезали на множество частей, почти все эти части погибали. Однако фрагменты, содержащие ядро, оказывались жизнеспособными и регенерировали все сложные клеточные структуры (рис. 1.5). Ядерный контроль клеточного морфогенеза и взаимодействие между ядром и цитоплазмой прекрасно иллюстрируют опыты на ацетабулярии (Acetabularia). Эта гигантская одиночная клетка (длиной в 2.5 - 5 см) состоит из трех частей: шляпки, стебелька и ризоида (рис. 1.6). Ризоид находится в основании клетки и прикрепляет ее к субстрату. Единственное клеточное ядро располагается в ризоиде. Большие размеры ацетабулярии и положение ее ядра позволяют исследователям удалить из клетки ядро и заменить его ядром другой клетки. В 30-х годах нашего века Геммерлинг (J. Hämmerling) воспользовался этими уникальными свойствами ацетабулярии и пересадил ядро одного вида (Acetabulariamediterranea) в энуклеированный ризоид другого (A. crenulata). Как видно из рис. 1.6. шляпки у этих видов очень сильно отличаются друг от друга. Геммерлинг обнаружил, что после того, как ядро одного вида трансплантировали в стебелек другого, новообразующаяся шляпка приобретала строение, свойственное
Гилберт С. Биология развития: В 3-х т. Т. I: Пер. с англ. — М.: Мир, 1993. — 228 с.ВВЕДЕНИЕ В ИЗУЧЕНИЕ РАЗВИТИЯ ЖИВОТНЫХ____________________________________________ 17
виду-донору ядра (рис. 1.7). Таким образом было показано, что развитие ацетабулярии контролируется ядром. Образование шляпки представляет собой сложный морфогенетический процесс, включающий синтез различных белков, которые должны распределиться между определенными частями клетки. Пересаженное ядро и в самом деле определяет образование видоспецифической шляпки, но для этого требуется несколько недель. Кроме того, если на ранней стадии развития ацетабулярии, до образования шляпки, удалить ядро, то через несколько недель образуется шляпка нормального строения, хотя организм в конце концов и погибнет. Это свидетельствует о том, что 1) ядро содержит информацию, определяющую тип образующейся шляпки (т.е. содержит генетическую информацию, которая обеспечивает синтез белков, ответственных за образование шляпки определенного типа), и 2) информация поступает в цитоплазму задолго до образования шляпки. Информация, поступившая в цитоплазму, не используется на протяжении нескольких недель. Одна из гипотез, предложенных для объяснения этих наблюдении, заключалась в том, что ядро синтезирует стабильную мРНК, которая остается в цитоплазме в неактивном состоянии, пока не наступит время образования шляпки. Более поздние данные, опирающиеся на наблюдение Геммерлинга (опубликованное в 1934 г.), подтверждают эту гипотезу. Геммерлинг разрезал молодую ацетабулярию на несколько частей (рис. 1.8). Шляпка развивалась не только на том фрагменте, в котором содержалось ядро (как этого и ожидали), но, кроме того, на апикальном конце стебелька. На средней части стебелька шляпка не развивалась. Исходя из этого, Геммерлинг постулировал (примерно за 30 лет до того, как узнали о существовании мРНК), что инструкции для образования шляпки каким-то образом сохраняются в концевой части стебелька. Результаты более новых исследований (Kloppstech, Schweiger. 1975) показали, что мРНК ядерного происхождения накапливаются именно в этой области. Прибавление к морской воде, в которой растет ацетабулярия, рибонуклеазы (фермента, расщепляющего РНК) полностью подавляет образование шляпки. В случае энуклеированных клеток это окончательный результат; коль скоро РНК разрушена, образования шляпки не происходит. Однако если в клетках ядро имеется, то после отмывания рибонуклеазы шляпка может обра- |