Билеты по микре. BILET_PO_MIKRE_ответы. Билет 1 Микробиология как фундаментальная наука. Ее задачи
 Скачать 0.54 Mb. Скачать 0.54 Mb.
|

|
3. Парамиксовирусы. Классификация, морфология. Диагностика. Характеристика заболеваний, вызванных этими вирусами Корь — острая инфекционная болезнь, характеризующаяся лихорадкой, катаральным воспалением слизистых оболочек верхних дыхательных путей и глаз, а также пятнисто-папулезной сыпью на коже. Таксономия. РНК-содержащий вирус. Семейства Paramyxoviridae. Род Morbillivirus. Структура и антигенные свойства. Вирион окружён оболочкой с гликопротеиновыми шипами. Под оболочкой находится спиральный нуклеокапсид. Геном вируса — однонитевая, нефрагменти-рованная минус РНК. Имеются следующие основные белки: NP — нуклеокапсидный; М — матриксный, а также поверхностные гли-козилированные белки липопротеиновой оболочки — гемагглютинин (Н) и белок слияния (F), гемолизин. Вирус обладает гемагглютинирующей и гемолитической активностью. Нейраминидаза отсутствует. Имеет общие антигены с вирусом чумы собак и крупного рогатого скота. Культивирование. Культивируют на первично-трипсинизированных культурах клеток почек обезьян и человека, перевиваемых культурах клеток HeLa, Vero. Возбудитель размножается с образованием гигантских многоядерных клеток — симпластов; появляются цитоплазматические и внутриядерные включения. Белок F вызывает слияние клеток. Резистентность. В окружающей среде нестоек, при комнатной температуре инактивируется через 3-4 ч. Быстро гибнет от солнечного света, УФ-лучей. Чувствителен к детергентам, дезинфектантам. Восприимчивость животных. Корь воспроизводится только на обезьянах, остальные животные маловосприимчивы. Эпидемиология. Корь — антропонозная инфекция, распространена повсеместно. Восприимчивость человека к вирусу кори чрезвычайно высока. Болеют люди разного возраста, но чаще дети 4—5 лет. Источник инфекции — больной человек. Основной путь инфицирования — воздушно-капельный, реже — контактный. Наибольшая заражаемость происходит в продромальном периоде и в 1-й день появления сыпи. Через 5 дней после появления сыпи больной не заразен. Патогенез. Возбудитель проникает через слизистые оболочки верхних дыхательных путей и глаз, откуда попадает в подслизистую оболочку, лимфатические узлы. После репродукции он поступает в кровь (вирусемия) и поражает эндотелий кровеносных капилляров, обуславливая тем самым появление сыпи. Развиваются отек и некротические изменения тканей. Клиника. Инкубационный период 8-15 дней. Вначале отмечаются острые респираторные проявления (ринит, фарингит, конъюнктивит, фотофобия, температура тела 39С). Затем, на 3—4-й день, на слизистых оболочках и коже появляется пятнисто-папулезная сыпь, распространяющаяся сверху вниз: сначала на лице, затем на туловище и конечностях. За сутки до появления сыпи на слизистой оболочке щек появляются мелкие пятна, окруженные красным ореолом. Заболевание длится 7—9 дней, сыпь исчезает, не оставляя следов. Возбудитель вызывает аллергию, подавляет активность Т-лимфоцитов и иммунные реакции, что способствует появлению осложнений в виде пневмоний, воспаления среднего уха и др. Редко развиваются энцефалит и ПСПЭ. Иммунитет. После перенесенной кори развивается гуморальный стойкий пожизненный иммунитет. Повторные заболевания редки. Пассивный иммунитет, передаваемый плоду через плаценту в виде IgG, защищает новорожденного в течение 6 месяцев после рождения. Микробиологическая диагностика. Исследуют смыв с носоглотки, соскобы с элементов сыпи, кровь, мочу. Вирус кори можно обнаружить в патологическом материале и в зараженных культурах клеток с помощью РИФ, РТГА и реакции нейтрализации. Характерно наличие многоядерных клеток и антигенов возбудителя в них. Для серологической диагностики применяют РСК, РТГА и реакцию нейтрализации. Лечение. Симптоматическое. Специфическая профилактика. Активную специфическую профилактику кори проводят подкожным введением детям первого года жизни или живой коревой вакцины из аттенуированных штаммов, или ассоциированной вакцины (против кори, паротита, краснухи). В очагах кори ослабленным детям вводят нормальный иммуноглобулин человека. Препарат эффективен при введении не позднее 7-го дня инкубационного периода. Билет 18 1. Микрофлора организма человека и ее роль в нормальных физиологических процессах и патологии. Микрофлора кишечника. 2. ГЗТ. Роль в противомикробном и противовирусном иммунитете. Аллергические пробы в лабораторной диагностике. 3. Вибриолы. Холера. Свойства: морфологические, культуральные, биохимические, антигенные. Факторы патогенности, токсины, специфическая прфилактика и терапия. Возбудитель – Vibrio cholerae, серогрупп О1 и О139, характеризуется токсическим поражением тонкого кишечника, нарушением водно-солевого баланса. Морфологические и культуральные свойства. Вибрион имеет один полярно расположенный жгутик. Под действием пенициллина образуются L-формы. Грамотрицательны, спор не образуют. Факультативный анаэроб. Не требователен к питательным средам. Температурный оптимум 37C. На плотных средах вибрионы образуют мелкие круглые прозрачные S-колонии с ровными краями. На скошенном агаре образуется желтоватый налет. В непрозрачных R-колониях бактерии становятся устойчивыми к действию бактериофагов, антибиотиков и не агглютинируются О-сыворотками. Биохимические свойства. Активны: сбраживают до кислоты глюкозу, мальтозу, сахарозу, маннит, лактозу, крахмал. Все вибрионы делятся на шесть групп по отношению к трем сахарам (манноза, сахароза, арабиноза). Первую группу, к которой относятся истинные возбудители холеры, составляют вибрионы, разлагающие маннозу и сахарозу и не разлагающие арабинозу: разлагают белки до аммиака и индола. H2S не образуют. Антигенная структура. Термостабильный О-антиген и термолабильный Н-антиген. Н-АГ являются общими для большой группы вибрионов. Возбудители классической холеры и холеры Эль-Тор объединяются в серогруппу 01. Антигены серогруппы 01 включают в различных сочетаниях А-, В- и С-субъединицы. Сочетание субъединиц АВ называется сероваром Огава, сочетание АС — сероваром Инаба, сочетание ABC — Гикошима. R-формы колоний утрачивают О-АГ. Резистентность. Вибрионы плохо переносят высушивание. Долго сохраняются в водоемах, пищевых продуктах.. Биовар Эль-Тор более устойчив в окружающей среде, чем классический вибрион. Эпидемиология. Острая кишечная инфекция с фекально-оральным механизмом передачи. Путь передачи - водный, пищевой. Источник инфекции — больной человек или вибрионоситель. Факторы патогенности. Пили адгезии; фермент муциназа, разжижающий слизь и обеспечивающий доступ к эпителию. Эпителиальные клетки выделяют щелочной секрет, который в сочетании с желчью является прекрасной питательной средой для размножения вибрионов. Токсинообразование вибрионов, которые вырабатывают эндо- и экзотоксины. Экзотоксин (энтеротоксин) холероген — термолабильный белок, чувствителен к протеолитическим ферментам. Холероген содержит 2 субъединицы: А и В. А активизирует внутриклеточную аденилатциклазу, происходит повышение выхода жидкости в просвет кишечника. Диарея, рвота. Фермент нейраминидаза усиливает связывание холерного экзотоксина с эпителием слизистой кишечника. Эндотоксин запускает каскад арахидоновой кислоты, которая запускает синтез простагландинов (Е, F). Они вызывают сокращение гладкой мускулатуры тонкого кишечника и подавляют иммунный ответ, чем обусловлены диарея. Клинические проявления. Инкубационный период 2—3 дня. Боль в животе, рвота, диарея. Иммунитет. Гуморально-клеточный. При выздоровлении возникает напряженный непродолжительный иммунитет. Микробиологическая диагностика. Выделение и идентификация возбудителя. Материал для исследования - выделения от больных (кал, рвота), вода. Для экспресс-диагностики используют РИФ, ПЦР. Бактериоскопический метод в настоящее время не используется. Лечение: а)регидратация (восполнение потерь жидкости и электролитов введением изотонических, растворов, а также плазмозаменяющих жидкостей внутривенно;б) антибактериальная терапия (тетрациклины, фторхинолоны). Профилактика. Санит.-гиг. мероприятия. Экстренная профилактика антибиотиками широкого спектра действия, а также вакцинопрофилактика. Современная вакцина представляет собой комплексный препарат, состоящий из холероген-анатоксина и химического О-антигена, обоих биоваров и сероваров Огава и Инаба. Прививка обеспечивает выработку вибриоцидных антител и антитоксинов в высоких титрах. Билет 19 1. Репродукция вирусов. Основные стадии взаимодействия вирусов с клеткой хозяев. Особенности репродукции ДНК- и РНК- содержащих вирусов. Генетика вирусов. Геном вирусов содержит или РНК, или ДНК (РНК- и ДНК- вирусы соответственно). Выделяют позитивную (+) РНК, обладающую матричной активностью и соответственно- инфекционными свойствами, и негативную ( - ) РНК, не проявляющую инфекционные свойства, которая для воспроизводства толжна транскрибироваться (превращаться) в +РНК. Механизмы репродукции различных вирусов очень сложные и существенно отличаются. Основные их схематические варианты представлены ниже. 1. вирионная (матричная) +РНК комплементарная -РНК (в рибосомах) вирионная +РНК. 2. - РНК вирусная (информационная) +РНК - РНК (формируется на геноме зараженной клетки). 3. однонитевая ДНК: +ДНК +ДНК -ДНК +ДНК -ДНК +ДНК +ДНК. 4. ретровирусная однонитевая РНК: РНК ДНК (провирус) РНК. 5. двунитевая ДНК: разделение нитей ДНК и формирование на каждой комплементарной нити ДНК. Генофонд вирусов создается и пополняется из четырех основных источников: двух внутренних (мутации, рекомбинации) и двух внешних (включение в геном генетического материала клетки хозяина, поток генов из других вирусных популяций). Комплементация- функциональное взаимодействие двух дефектных вирусов, способствующее их репликации и горизонтальной передаче. Фенотипическое смешивание- при заражении клетки близкородственными вирусами с образованием вирионов с гибридными капсидами, кодируемыми геномами двух вирусов. Популяционная изменчивость вирусов связана с двумя разнонаправленными процессами - мутациями и селекцией, связанными с внешней средой как индуктором мутаций и фактором стабилизирующего отбора. Гетерогенность вирусных популяций- адаптационный генетический механизм, способствующий пластичности (устойчивости, приспособляемости) популяций, фактор эволюции и сохранения видов во внешней среде. Генофонд вирусных популяций сохраняется за счет нескольких механизмов: - восстановления изменчивости за счет мутаций; - резервирующих механизмов (возможность перехода любых, даже негативных мутаций в следующую генерацию)- комплементация, рекомбинация; - буферных механизмов (образование дефектных вирусных частиц, иммунных комплексов и др.), способствующие сохранению вируса в изменяющихся внешних условиях. Типы взаимодействия вируса с клеткой. Стадии репродукции вирусов. Типы взаимодействия вируса с клеткой. Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный. Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма). Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов. Интегративный тип, или вирогения — характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация). Репродукция вирусов осуществляется в несколько стадий, последовательно сменяющих друг друга: адсорбция вируса на клетке; проникновение вируса в клетку; «раздевание» вируса; биосинтез вирусных компонентов в клетке; формирование вирусов; выход вирусов из клетки. Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 104 до 105. Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц. Проникновение в клетку. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга. «Раздевание». Процесс «раздевания» заключается в удалении защитных вирусных оболочек и освобождении внутреннего компонента вируса, способного вызвать инфекционный процесс. «Раздевание» вирусов происходит постепенно, в несколько этапов, в определенных участках цитоплазмы или ядра клетки, для чего клетка использует набор специальных ферментов. В случае проникновения вируса путем слияния вирусной оболочки с клеточной мембраной процесс проникновения вируса в клетку сочетается с первым этапом его «раздевания». Конечными продуктами «раздевания» являются сердцевина, нуклеокапсид или нуклеиновая кислота вируса. Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства. Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации. Формирование (сборка) вирусов. Синтезированные вирусные нуклеиновые кислоты и белки обладают способностью специфически «узнавать» друг друга и при достаточной их концентрации самопроизвольно соединяются в результате гидрофобных, солевых и водородных связей. Существуют следующие общие принципы сборки вирусов, имеющих разную структуру: 1. Формирование вирусов является многоступенчатым процессом с образованием промежуточных форм; 2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа); 3. Формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки; 4. Сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы). Выход вирусов из клетки. Различают два основных типа выхода вирусного потомства из клетки. Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки. Второй тип — почкование. Он присущ вирусам, имеющим суперкапсидную оболочку. На заключительном этапе сборки нуклеокапсиды сложно устроенных вирусов фиксируются на клеточной плазматической мембране, модифицированной вирусными белками, и постепенно выпячивают ее. В результате выпячивания образуется «почка», содержащая нуклеокапсид. Затем «почка» отделяется от клетки. Таким образом, внешняя оболочка этих вирусов формируется в процессе их выхода из клетки. При таком механизме клетка может продолжительное время продуцировать вирус, сохраняя в той или иной мере свои основные функции. Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции. 2. Антитела. Классификация иммуноглобулинов. Динамика антителообразования. Следовательно, антитела — это иммуноглобулины, вырабатываемые в ответ на введение антигена и способные специфически взаимодействовать с этим же антигеном. Функции. Первичная функция состоит во взаимодсйствии их активных центров с комплементарными им детерминантами антигенов. Вторичная функция состоит в их способности:Нейтрализация токсинов,лизис бактерий.при.участии.комплимента,опсонизация и усиление фагацитоза,участвовать в распознавании «чужого» антигена и его связывание;нейтрализация внеклеточ вируса,цитотоксическое действие на вирус, обеспечивать кооперацию.иммунокомпетентных клеток (макрофагов, Т- и В-лимфоцитов);участвовать в различных формах иммунного ответа . Иммуноглобулины по структуре, антигенным и иммунобиологическим свойствам разделяются на пять классов: IgM-5-10% в крови,молек масса 900 000,5 мономеров, валентность 10,содерж.в сыворотке крови- 1г/л, синтезируется В-лимфоцитами,не проходит через плаценту,функции:первич.имун.ответ,может образовывать секреторную форму и выделяться в молоко,обеспечивает нейтрализацию,опсонизацию и маркирование антигена,осуществляет запуск комплимент-опосредованного цитолиза и антителозависимой клеточно-опосредованной цитотоксичности. IgG-75-85%ур.крови,150 000 масса,1 мономер,валентность2, в сыворотке 12г/л,проходит через плаценту и обеспеч гуморал имунитет новорожденного в первые 3 месяца,участвует во 2-ом имун.ответе, синтезируется В-лимфацитами и плазматическими клетками,участвует в развитии аллерг.реакции 1-го типа, выделяется в секрет слизистыхи в молоко путем диффузии. IgA-7-5% в крови молек.масса 170 000(сывороточный) или 350 000(секреторный), Сывороточный- 1 мономер,валентность 2,синтез. В-лимфоцитами и плазматич.клетками.Облад.высок. афинностью,не связ комплемент,не проходит через плацентарный барьер.Секреторный-2 мономера,валентность 4,синтез.В-лимфоцитами,Секретор Ig-основной фактор спецефического гуморалного местного иммунитета слизистых оболочек ЖКТ,мочеполовой системы и респираторного тракта,он препятствует адгезии микробов на эпителиальных клетках и генерализации инфекции в пределах слизистых. IgE-0,002%в крови,масса 190 000,1 мономер, валентность 2,содерж в сыворотке 0,00025 г/л,синтезируется В-лимфацитами, плазматическими клетками в лимфоидной ткани,бронхолегочной ткани ,ЖКТ, не связ.комплемент, не проходит через плаценту,является реагином-защита от паразитов. IgD-1%в крови,масса 180 000,мономеры 1, валентость 2,в сыворотке 0,03г/л,является мембранным рецептором.. Молекулы иммуноглобулинов всех пяти классов состоят из полипептидных цепей: двух одинаковых тяжелых цепей Н и двух одинаковых легких цепей — L, соединенных между собой дисульфидными мостиками. Как в Н-, так и в L-цепях имеется вариабельная — V область, в которой последовательность аминокислот непостоянна, и константная — С область с постоянным набором аминокислот. В легких и тяжелых цепях различают NH2- и СООН-концевые группы. При воздействии протеолитическим ферментом папаином иммуноглобулин расщепляется на три фрагмента: два не кристаллизующихся, содержащих детерминантные группы к антигену и названных Fab-фрагментами I и II и один кристаллизующий Fc-фрагмент. FabI- и FabII-фрагменты сходны по свойствам и аминокислотному составу и отличаются от Fc-фрагмента; Fab-и Fc-фрагменты являются компактными образованиями, соединенными между собой гибкими участками Н-цепи, благодаря чему молекулы иммуноглобулина имеют гибкую структуру. Как Н-цепи, так и L-цепи имеют отдельные, линейно связанные компактные участки, названные доменами; в Н-цепи их по 4, а в L-цепи — по 2. Активные центры, или детерминанты, которые формируются в V-областях, занимают примерно 2 % поверхности молекулы иммуноглобулина. В каждой молекуле имеются две детерминанты, относящиеся к гипервариабельным участкам Н-и L-цепей, т. е. каждая молекула иммуноглобулина может связать две молекулы антигена. Поэтому антитела являются двухвалентными. 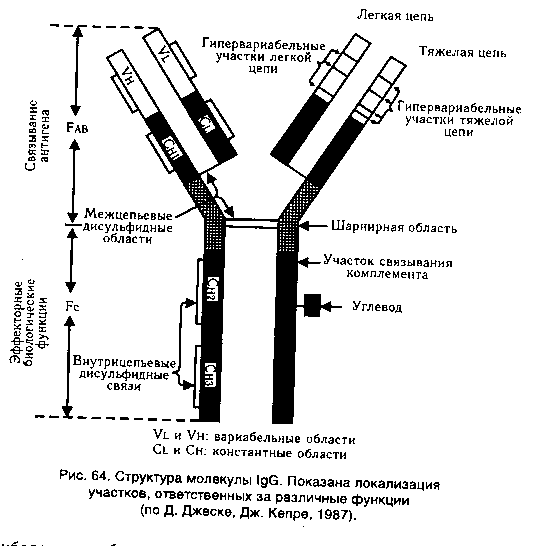 |